
- Search this journal
- Search all journals

Article Sections
- Materials and Methods
- Results and Discussion
Article Figures
Vegetative top dry weight (DW), leaf DW, stem DW, and stem diameter of mature eggplant plants as affected by irrigation rate. Irrigation rate was applied as percentage of crop evapotranspiration. Curve was fit by linear regression. Fall of 2010, Tifton, GA.
Seasonal volumetric soil water content (measured at 12- and 30-cm depth) as influenced by irrigation rate. Irrigation rate was applied as percentage of crop evapotranspiration. Line was fit by linear regression. Fall of 2010, Tifton, GA.
Effect of irrigation rate and soil depth on the concentration of nitrate-nitrogen in the soil (0 to 60 cm) in drip-irrigated eggplant grown on raised beds and plastic film mulch. Irrigation rate was applied as percentage of crop evapotranspiration. Line was fit by linear regression. Fall of 2010, Tifton, GA.
Cumulative number of fruit and fruit yields as affected by irrigation rate in drip-irrigated eggplant grown on raised beds and plastic film mulch. Irrigation rate was applied as percentage of crop evapotranspiration. Line was fit by linear regression. Fall of 2010, Tifton, GA.
Individual fruit weight as influenced by irrigation rate in drip-irrigated eggplant grown on raised beds and plastic film mulch. Irrigation rate was applied as percentage of crop evapotranspiration. Line was fit by linear regression. Fall of 2010, Tifton, GA.
- View raw image
- Download Powerpoint Slide
Related Content
- Previous Article
- Next Article
Eggplant ( Solanum melongena L.) Plant Growth and Fruit Yield as Affected by Drip Irrigation Rate
Click on author name to view affiliation information
Eggplant ( Solanum melongena L.) is an increasingly popular crop in the United States. In the southeastern United States, eggplant is often produced with high levels of irrigation water [above the rate of crop evapotranspiration (ETc)], resulting in water waste and nitrogen (N) leaching. The objective of this research was to assess the effects of irrigation rate on plant growth and fruit yield in eggplant. The study was conducted in Tifton, GA, in the fall of 2010 and 2011. Eggplant plants cv. Santana were grown on raised beds (1.8 m centers) covered with white plastic film mulch. There was a single drip tape along the center of the bed. The design was a randomized complete block with five treatments and four replications. Treatments consisted of irrigation rates based on ETc (33%, 67%, 100%, 133%, and 167% ETc). Plant growth, chlorophyll index (CI), and volumetric soil water content (SWC) were monitored over the season. In 2010, SWC (0–30 cm deep) increased and soil nitrate levels decreased with increasing irrigation rates. Foliar N and potassium (K), and CI decreased with increasing irrigation rate, probably due to a dilution effect. Stem diameter, leaf dry weight (DW), and vegetative top DW increased with increasing irrigation rate. Net photosynthesis and stomatal conductance ( g S ) were lowest at 33% ETc. Fruit number and fruit yields (marketable and total) were also lowest at 33% ETc and there were little yield differences among irrigation rates higher than 33% ETc. In 2011, irrigation rate had minor or no effect on SWC, plant growth of mature plants, leaf gas exchange, and fruit number and yield. The no treatment effect observed for eggplant in 2011 was likely because study was conducted in a low field that remained moist most of the time, nullifying the treatment effects. Results suggested that eggplant may tolerate mild water stress, since plants irrigated at 67% ETc produced fruit yields similar to those of plants irrigated at 100% ETc or higher rates. Thus, there is a potential to save water by reducing current irrigation rates without negatively impacting fruit yields.
Eggplant, also known as aubergine and brinjal, is widely grown and consumed in southern and southeast Asia and has increased in popularity in the United States as a specialty vegetable. In 2001, U.S. eggplant production was valued at $42.5 million, and Georgia, Florida, California, New Jersey, and New York were the top five producers. The U.S. Department of Agriculture has not collected complete domestic production statistics for eggplants since 2001. In 2012, farm gate value in the state of Georgia was $17 million ( CAED, 2013 ). Average eggplant yield in Florida is ≈30 t·ha −1 ( Ozores-Hampton, 2014 ).
Eggplant is in the Solanaceae family, as are tomato ( Solanum lycopersicon ) and pepper ( Capsicum annum ) and shares similar environmental and cultural requirements as those crops. However, in contrast to tomato and pepper, eggplant crop can tolerate greater levels of drought stress ( Behboudian, 1977 ). There are several studies on eggplant irrigation carried out in Asia, Africa, and Europe ( Aujla et al., 2007 ; Behboudian, 1977 ; Chartzoulakis and Drosos, 1995 ; Gaveh et al., 2011 ; Karam et al., 2011 ) showing that eggplant can be produced at moderate levels of drought stress without major impact on fruit yield.
In southeastern United States, eggplant is often produced with high levels of irrigation water (above the rate of ETc) and N fertilizer, resulting in water waste and N leaching. Excessive irrigation rate not only wastes water, but may also result in reduced yields in bell pepper ( Díaz-Pérez et al., 2004 ; Sezen et al., 2006 ) and tomato ( Locascio et al., 1989 ; Ngouajio et al., 2007 ). To our knowledge, there are no published studies in the United States on the effect of irrigation rate on the yield and plant growth of drip-irrigated eggplants. Irrigation studies, intended to optimize use of irrigation water, are necessary to enable the protection of water resources in the United States. Therefore, the objective of this research was to assess the effects of irrigation rate on plant growth and fruit yield in eggplant.
Study site.
The study was carried out at the Horticulture Farm, University of Georgia, Tifton, GA, during the fall of 2010 and 2011. The farm is located at an altitude of 108 m above mean sea level, 31°28′ N latitude and 83°31′ W longitude. The soil of the farm is a Tifton sandy loam (a fine loamy-siliceous, thermic Plinthic Kandiudults) with pH 6.5. Available water capacity is 18 to 36 mm in the top 30 cm of soil profile ( Calhoun, 1983 ). In 2010, field had a gentle sloping (slope ≈3%); in 2011, field had a nearly level slope. The distance between the 2010 and 2011 fields was ≈70 m.
Land preparation and planting.
Eggplant plants were grown on plastic film mulch on raised beds (6 × 0.76 m, formed on 1.8-m centers). Before laying mulch, the soil was fertilized with N, phosphorous (P), and K at 60, 26, and 50 kg·ha −1 , respectively, using 10–10–10 granular fertilizer. At the same time, plastic film mulch [white on black, low-density polyethylene with a slick surface texture, 1.52 m wide and 25 µm thick (RepelGro, ReflecTek Foils, Inc., Lake Zurich, IL)] was laid with a mulch-laying machine, drip irrigation tape [20.3 cm emitter spacing and a 8.3 mL·min −1 emitter flow (Ro-Drip, Roberts Irrigation Products, Inc., San Marcos, CA)] was placed 5 cm deep in the center of the bed.
Eggplant transplants were produced in a greenhouse using peat-based medium (Pro-Mix, Quakertown, PA) and polystyrene 200-cell (2.5 × 2.5 cm cell) trays. Six-week-old eggplant transplants were planted with a mechanical transplanter on 6 Aug. 2010 and 5 Aug. 2011 in one row per bed, with a 60 cm separation between plants. About 250 mL of starter fertilizer solution (555 mg·L −1 N; 821·mg·L −1 P; 0 mg·L −1 K) was applied directly to the base of each transplant. The length of the experimental plot was 6.1 m. Starting 3 weeks after transplanting, plants were fertilized weekly through the drip system with N and K. Fertilization rates of N and K after transplanting were 0.7, 1.0, 1.5, and 2 kg·ha −1 ·d −1 in week 5, week 6; week 7; and weeks 13–15, respectively. Total N–P–K applied in the season was 218 kg·ha −1 N, 30 kg·ha −1 P, and 181 kg·ha −1 K.
Experimental design and treatments.
The design was a randomized complete block with five treatments and four replications. Treatments consisted of irrigation rates based on ETc (33%, 67%, 100%, 133%, and 167% the rate of ETc). ETc was calculated by multiplying the reference evapotranspiration (ETo) by a crop coefficient (Kc), which is dependent on the crop stage of development. Available Kc values for eggplant were developed for bare soil (unmulched) production. These Kc values, however, are not recommended for crops under plasticulture systems since plastic mulches reduce soil evaporation and ETc ( Allen et al., 1998 ; Pereira et al., 2015 ; Simonne et al., 2006 ). The Kc values used in this study were modified relative to those proposed for bell pepper in Florida ( Simonne et al., 2006 ). The Kc values used were 0.25 (week 1 after transplanting), 0.40 (week 2), 0.55 (week 3), 0.70 (week 4), 0.85 (week 5), 1.0 (week 6–11), and 0.8 (week 12–14).
All treatments received equal volumes of irrigation water (88 and 49 mm in 2010 and 2011, respectively) during the crop establishment period (first 4 weeks after transplanting). Irrigation treatments were initiated on week 5. Water was applied when cumulative ETc was ≈12 mm, which corresponded to about every 2 to 3 d in mature plants (mean ETo was 5 to 6 mm·d −1 ). Thus, amounts of water per irrigation event were ≈4 mm (33% ETc), 8 mm (67% ETc), 12 mm (100% ETc), 16 mm (133% ETc), and 20 mm (167% ETc).
Soil water content.
Soil water content (volumetric) in the 0–12 cm of soil profile over the season was measured manually once every 2–3 d (three readings per experimental plot) with a portable time-domain reflectometry (TDR) sensor (CS-620; Campbell Scientific, Logan, UT). The two metallic 12-cm rods of the TDR sensor were inserted vertically within the row between two plants. Soil water content (volumetric) in the 0–30 cm of soil profile was periodically (every 10 min) monitored with TDR sensors (CS-610; Campbell Scientific) connected to a datalogger (CR-10X; Campbell Scientific). The moisture sensors had three metallic 30-cm rods and were inserted vertically within the row between two plants.
Soil nitrate.
Soil samples were taken from each plot at 0- to 20-cm, 20- to 40-cm, and 40- to 60-cm depths on 8 Nov. 2010. Samples were taken at least 0.5 m away from the borders of the plots and from the previous sampling holes. Samples were air-dried and analyzed for nitrate-nitrogen using standard QuickChem Methods (Lachat Quick-Chem 8000 FIA; Zellweger Analytics, Milwaukee, WI).
Plant growth.
Eggplant plant height and stem diameter were measured weekly in three mature plants per plot. Plant samples obtained at the end of the season were dried at 70 °C for several days until constant weight was obtained. Leaf, stem, and vegetative top (leaf + stem) DW of individual plants were determined.
Chlorophyll indices were determined twice a week over the season on six mature, well-exposed, and healthy leaves per plot using a chlorophyll meter (Chlorophyll Meter SPAD-502; Minolta Co., Ltd., Ramsey, NJ).
Leaf gas exchange and PSII efficiency.
Simultaneous measurements of leaf gas exchange (net photosynthesis, g S , transpiration, and internal CO 2 concentration), and fluorescence were determined as PSII efficiency were made with an infrared gas analyzer (LI-COR 6400 IRGA with an integrated 6400-40 leaf chamber fluorometer; LI-COR, Inc., Lincoln, NE). PSII efficiency is the fraction of absorbed PSII photons used in photochemistry and is measured with a light-adapted leaf. Water use efficiency (WUE) was calculated as the ratio between leaf net photosynthesis and leaf transpiration. Air flow rate was set at 300 µmol·m −2 ·s −1 on the reference side. The CO 2 concentration was set at 400 µmol·mol −1 with a CO 2 mixer and a CO 2 tank. Measurements were conducted in developed plants on clear days (photosynthetically active radiation ≈2000 µmol·m −2 ·s −1 ) at 1200–1500 hr Eastern Standard Time in 2010 (6 and 20 Oct. and 9 Nov.) and 2011 (5 Oct.), using two developed and fully exposed leaves per experimental plot.
Leaf mineral nutrients.
Leaf samples (20 fully developed leaves from new growth) from developed plants were dried at 70 °C for 2 d and analyzed for mineral nutrient concentration at the University of Georgia, Agricultural & Environmental Services Laboratories, Athens, GA.
Weather data (air temperature, ETo, and rainfall) were obtained from a nearby University of Georgia weather station (within 300 m).
The harvest lasted from 28 Sept. to 23 Nov. in 2010 and from 23 Sept. to 4 Nov. in 2011. Eggplant fruit were harvested twice per week at commercial stage. Harvested section consisted of 10 plants per plot. Fruit were graded according to U.S. Department of Agriculture standards ( USDA, 2013 ) as marketable or cull and number and weight of marketable and cull fruit were determined. Average fruit weight was derived mathematically from the total weight and the total number of fruits.
Irrigation water use efficiency.
Irrigation water use efficiency (IWUE) was calculated by dividing fruit weight (kg·ha −1 ) by irrigation water received by the crop (in mm) for each irrigation treatment.
Agronomic efficiency of nitrogen.
Agronomic efficiency of nitrogen was calculated by dividing total eggplant fresh fruit weight (kg·ha −1 ) by the amount of N (kg·ha −1 ) applied to the crop.
Fruit DW content and harvest index (HI).

Statistical analysis.
Data were analyzed using the General Linear Model and Regression Procedures from SAS (SAS version 9.3, SAS Institute Inc., Cary, NC). Data means were separated by Fisher’s protected least significant difference test at 95% confidence and response curves determined by orthogonal contrasts. Percentages were transformed to arcsin values before analysis. For clarity, nontransformed percentage means were used for presentation in tables and figures. Data from all years were pooled if no year × treatment interactions were found.
In 2010, average maximal, mean, and average minimum air temperature for the season were 28.8, 22.6, and 16.4 °C, respectively. Cumulative ETo and rainfall for the season were 370 and 184 mm, respectively. In 2011, average maximal, mean, and average minimum air temperature were 28.6, 22.5, and 16.4 °C, respectively. Cumulative ETo and rainfall for the season were 344 and 256 mm, respectively.
In 2010, vegetative top DW, leaf DW, stem DW, and stem diameter increased with increasing irrigation rate ( Fig. 1 ). Leaf weight ratio (LWR) [leaf biomass as a fraction of vegetative aboveground biomass (mean = 0.529)] decreased with increasing irrigation rate ( r 2 = 0.92; P ≤ 0.05) from LWR of 0.543 at 33% ETc to LWR of 0.493 at 167% ETc, which indicates that plants allocated less biomass to leaves as irrigation rate increased. Bell pepper leaves have reduced leaf thickness at low light and low water stress conditions ( Díaz-Pérez, 2013 ). In 2011, over the season, mean stem diameter was lowest at 33% ETc ( P < 0.05), although final stem diameter was unaffected by irrigation rate ( Table 1 ). Mean seasonal plant height increased with irrigation rate, ranging from 66 cm (33% ETc) to 93 cm (167% ETc); final plant height (4 Nov.) was unaffected by irrigation rate. Mature plant DW (mean = 1.70 kg) was also unaffected by irrigation rate. Growth differences during midseason but not at the end of the season were probably because of high evaporative demand conditions that impacted plant growth at low irrigation rates during midseason. Late in the season, when evaporative demand was reduced, the effect of irrigation rate on plant growth was less detectable. The no treatment effect observed for eggplant in 2011 was likely because study was conducted in a low field that remained moist most of the time, nullifying the treatment effects.

Citation: HortScience horts 50, 11; 10.21273/HORTSCI.50.11.1709
- Download Figure
Plant growth, leaf chlorophyll index (CI), and soil water content (SWC) as affected by irrigation rate in eggplant. Fall of 2011, Tifton, GA.

Reduced eggplant plant growth at irrigation rates below 100% ETo has been previously reported. Eggplant irrigated at 80% pan evaporation, every 8 d, and 70% pan evaporation, every 12 d, had reduction of 18% and 27% in plant height, and 13% and 21% in stem diameter, respectively ( Kirnak et al., 2002 ). In bell pepper exposed to different soil water levels by varying drip emitter spacing, plant height and canopy diameter increased with decreasing emitter spacing (i.e., with increased soil water levels) ( Madramootoo and Rigby, 1991 ).
In 2010, CIs decreased with increased irrigation rate ( P = 0.006), from 60.8 at 33% ETc to 59.0 at 167% ETc. In 2011, CI decreased from 55.8 at 33% ETc to 53.7 at 167% ETc ( Table 1 ). Decreased CI values with increased irrigation rates were likely due to dilution effect of nutrients, since plant growth was enhanced with increased irrigation rates. Decreased CI with increased irrigation rates may also be associated with increased nitrate leaching under high irrigation rates.
In 2010, the effect of irrigation rate on SWC varied with soil depth. At 0- to 30-cm depth, SWC increased with increasing irrigation rates ( Fig. 2 ), whereas at 0- to 12-cm depth SWC was unaffected by irrigation rate. Differences in soil moisture in the different soil depths indicate a higher soil water uptake by plants, because of greater presence of roots at 0–12 cm than at 0- to 30-cm depth; they also indicate that high rates of irrigation (>100% ETc) result in wasted water because much water at 0- to 30-cm depth was not taken up by the crop; and they suggest that soil moisture measurement at 0- to 30-cm depth was more sensitive to detect changes in soil moisture than measurement at 0- to 12-cm depth.

As in 2010, seasonal SWC at 0- to 12-cm depth was also similar among irrigation rates (mean = 13.4%) in 2011. In addition to the high presence of roots at 0- to 12-cm depth, SWC values were similar among treatments in 2011 probably because the study was conducted in a low field, with a nearly level slope, where soil was commonly moist throughout the season, likely due to lateral water movement from upper sections of the field. There was an impermeable clay layer 30- to 40-cm deep in the soil profile that probably allowed water to flow from upper to lower areas within the farm.
Leaf gas exchange.
In 2010, the effect of irrigation rate on leaf gas exchange varied by date ( Table 2 ). Net photosynthesis, g S , and photosynthetic WUE were unaffected by irrigation rate on 6 Oct. 2010. Lack of treatment differences on 6 Oct. was probably attributable to relatively low temperatures on day of measurement (mean temperature = 16.4 °C), resulting in low crop evaporative demand and low crop water stress. Net photosynthesis and g S were lowest at 33% ETc on 20 Oct. and 9 Nov. Water use efficiency was highest and PSII efficiency was lowest at 33% ETc on 20 Oct. The fact that gas exchange variables were not reduced at 67% ETc compared with higher irrigation rates suggests that plants at 67% ETc were likely unaffected by water stress. However, since gas exchange measurements were conducted only in mature plants, late in the season, when evaporative demand was reduced, it is possible that earlier in the season plants may have had experienced increased water stress at reduced irrigation rates, as suggested by the reduced plant growth at reduced irrigation rates. In 2011, leaf net photosynthesis (mean = 28.3 µmol·m −2 ·s −1 ), g S (mean = 0.248 mol·m −2 ·s −1 ), WUE (mean = 4.24 µmol·mmol −1 ), and PSII (mean = 0.189 µmol·mmol −1 ) were unaffected by irrigation rate. Air maximal and minimal temperature on the day of measurement were 27.5 and 11.0 °C, respectively. Lack of differences in gas exchange are consistent with the lack of differences in plant growth among irrigation rates observed in 2011.
Leaf gas exchange and fluorescence as affected by irrigation rate and date in eggplant. Fall of 2010, Tifton, GA.

Irrigation at 33% ETc was probably insufficient to satisfy eggplant water requirements, as suggested by the reduced leaf gas exchange values ( Table 2 ). Reduced irrigation rates can result in decreased gas exchange in solanaceous crops. Transpiration, leaf g S , and leaf net photosynthesis in eggplant were reduced with water stress and effects varied depending on stress severity and duration ( Sarker et al., 2005 ). In habanero pepper ( Capsicum chinense Jacq.), there was reduced g S and net photosynthesis with increased time between irrigations ( Jaimez et al., 1999 ).
Soil nitrate concentration decreased with increasing irrigation rate ( P = 0.002) and soil depth ( P = 0.003), indicating that nitrate leaching to the deepest parts of the soil was enhanced with increased irrigation rates ( Fig. 3 ). Decreased soil nitrate concentration may also be due to high N uptake by the crop, as suggested by augmented vegetative growth with increasing irrigation rate. Nitrate present at 40–60 depth is usually lost as it is not recovered by plants’ roots. Decreased nitrate in 40- to 60-cm zone is thus solely due to leaching.

Foliar mineral nutrient concentrations and CI.
In 2010, foliar N and K concentrations decreased and P increased with increasing irrigation rate ( Table 3 ). Other foliar nutrients concentrations were unaffected by irrigation rate. Nitrogen, K, and CI decreased with irrigation rate, possibly as a result of a dilution effect associated with increased aboveground plant growth. In addition, at high irrigation rates plants likely had reduced access to soil N due to increased nitrate leaching. Plant water stress in eggplant can reduce foliar N, P, and K concentrations compared with well-irrigated plants ( Kirnak et al., 2002 ). In the present study, however, only foliar P was reduced at low irrigation rate.
Foliar mineral nutrient concentrations in eggplant as affected by several irrigation rates. Fall of 2010, Tifton, GA. z

Chlorophyll indices have been used as indirect estimators of chlorophyll and leaf N concentrations ( Liu et al., 2006 ). Crop drought stress may influence leaf morphology (e.g., increased specific leaf weight) in plants ( Larcher, 1995 ); these variations in leaf morphology may also influence CI, making difficult to use CI to estimate leaf N ( Díaz-Pérez, 2013 ). In our study, CI values increased with increasing leaf N ( R 2 = 0.921; P = 0.001), supporting the use of chlorophyll meter to estimate leaf N.
In 2010, fruit number and fruit yields (marketable and total) were lowest at 33% ETc and there were little yield differences among irrigation rates higher than 33% ETc ( Fig. 4 ). Individual fruit weight was also reduced at 33% ETc ( Fig. 5 ). There was a higher correlation between fruit number and fruit yield ( R 2 = 0.94; P < 0.0001) than between individual fruit weight and fruit yield ( R 2 = 0.15; P = 0.027), suggesting that marketable yield was determined more by fruit number than individual fruit weight. In greenhouse-grown eggplant, soil water deficit decreased fruit number but not fruit size ( Chartzoulakis and Drosos, 1995 ). In a study with different levels of irrigation and N fertilizer, eggplant fruit yield was more related with fruit number than with fruit size ( Aujla et al., 2007 ). In another study, soil water deficits also reduced eggplant fruit size, but the effect of drought stress on fruit number was not evaluated ( Kirnak et al., 2002 ). In 2011, irrigation rate had no effect on the number or yields of marketable, cull, and total fruit, or on individual fruit weight ( Table 4 ). There were no significant interactions between harvest dates and irrigation rates. There was also a higher correlation between fruit number and fruit yield ( R 2 = 0.92; P < 0.0001) than between individual fruit weight and fruit yield ( R 2 = 0.185; P = 0.001). Results suggest that eggplant may tolerate moderate water stress, since plants irrigated at 67% ETc produced fruit yields similar to those of plants irrigated at 100% ETc or higher rates. Thus, there is a potential to reduce irrigation rates below 100% ETc without negatively impacting fruit yields.

Fruit yield of eggplant as affected by irrigation rate. Fall of 2011, Tifton, GA.

Irrigation water use efficiency and agronomic efficiency of nitrogen.
Plants received more irrigation water in 2010 than in 2011 as a result of reduced rainfall in 2010 ( Table 5 ). In both years, IWUE decreased with increasing irrigation rate. IWUE was greatly reduced and there were significant effects of irrigation rates on several variables in 2010, but not in 2011. Increased IWUE and increased SWC in 2011 (mean = 13.4% at 0- to 12-cm depth) relative to SWC in 2010 (mean = 7.5% at 0- to 12-cm depth) are probably associated with increased contribution of soil water from rainfall and drainage water from upper areas of the field; in 2011, field used was low and nearly flat.
Irrigation, cumulative rainfall, IWUE, and AEN of eggplant crop grown on plastic film mulch. Fall of 2010 and 2011, Tifton, GA.

Although there were differences in leaf N among irrigation treatments, fruit yield was likely more related to irrigation rate than to leaf N. Total yield showed a quadratic relationship with leaf N ( R 2 = 0.185; P = 0.013); total yield was unaffected by leaf N below 5.1% and was lowest at the highest leaf N (5.3%) occurred at the lowest irrigation rate (33% ETc).
Agronomic efficiency of N increased with irrigation rate in 2010 likely as a result of increased fruit yield associated with improved plant water status; AEN was unaffected by irrigation rate in 2011. AEN values in this study (range 92 to 187 kg·kg −1 N) were lower compared with values of other studies on eggplant (range = 324 to 859 kg·kg −1 N) ( Aujla et al., 2007 ), probably because the harvest period in this study was reduced. Low AEN values may also mean that eggplant crop in this study made inefficient use of N fertilizer, probably in part due to overfertilization. Aujla et al. (2007) reported that irrigation rate and N fertilization rate interacted in drip-irrigated eggplants; they also found that irrigation at 75% pan evaporation and 120 kg·ha −1 N produced the greatest yields, and that AEN increased with increased N fertilization rate.
Fruit DW content and HI.
In year 2010, fruit DW content (mean = 6.2%) was unaffected by irrigation rate. In a study under semiarid conditions, soluble DW or soluble solids in eggplant decreased with increased irrigation rates ( Kirnak et al., 2002 ). In greenhouse-grown eggplant, increased irrigation rates also decreased fruit DW content ( Chartzoulakis and Drosos, 1995 ).
Harvest index was unaffected by irrigation rate (mean HI = 0.32). These data suggest that eggplant is more tolerant to drought than other solanaceous crops ( Behboudian, 1977 ). Our measurements of HI did not include root biomass. However, under water stress, eggplants possibly allocated increased amounts of assimilates for root growth as occurs in other plants ( Larcher, 1995 ). In habanero pepper, an irrigation rate of 20% of available water produced reduced values of HI ( Quintal Ortiz et al., 2012 ). In tomato, there was no difference in total dry biomass and HI between the control and a partial irrigation treatment, but total dry biomass and HI significantly decreased under regulated deficit irrigation ( Lei et al., 2009 ); moderate water stress–induced osmotic regulation under partial root drying conditions, leading to normal water status and the same level of biomass. Eggplant in our study was also able to maintain high fruit yields at moderate levels of water stress, suggesting that, as tomato, eggplant is also able to develop mechanisms to deal with water stress such as osmoregulation.
In conclusion, the results from this research indicate that eggplant may tolerate moderate water stress, since plants irrigated at 67% ETc had no detrimental effects on plant growth and leaf gas exchange and produced fruit yields similar to those of plants irrigated at 100% ETc. Thus, there is a potential to reduce current irrigation rates without negatively impacting fruit yields or quality.
Literature Cited
Allen, R.G. , Pereira, L.S. , Raes, D. & Smith, M. 1998 Crop evapotranspiration: Guidelines for computing crop water requirements. Food and Agriculture Organization of the United Nations, Rome
Aujla, M.S. , Thind, H.S. & Buttar, G.S. 2007 Fruit yield and water use efficiency of eggplant ( Solanum melongema L.) as influenced by different quantities of nitrogen and water applied through drip and furrow irrigation Sci. Hort. 112 142 148
- Search Google Scholar
- Export Citation
Behboudian, M.H. 1977 Responses of eggplant to drought. I. Plant water balance Sci. Hort. 7 303 310
CAED 2013 2012 Georgia farm gate value report. CAES, Univ. of Georgia, Athens, GA
Calhoun, J.W. 1983 Soil survey of Tift County, Georgia. United States Department of Agriculture, Soil Conservation Service, University of Georgia, GA
Chartzoulakis, K. & Drosos, N. 1995 Water use and yield of greenhouse grown eggplant under drip irrigation Agr. Water Mgt. 28 113 120
Díaz-Pérez, J.C. 2013 Bell pepper ( Capsicum annum L.) crop as affected by shade level: Microenvironment, plant growth, leaf gas exchange, and leaf mineral nutrient concentration HortScience 48 175 182
Díaz-Pérez, J.C. , Granberry, D. , Seebold, K. , Giddings, D. & Bertrand, D. 2004 Irrigation levels affect plant growth and fruit yield of drip-irrigated bell pepper HortScience 39 748
Gaveh, E.A. , Timpo, G.M. , Agodzo, S.K. & Shin, D.H. 2011 Effect of Irrigation, transplant age and season on growth, yield and irrigation water use efficiency of the African eggplant J. Hort. Environ. Biotechnol. 52 13 28
Jaimez, R.E. , Rada, F. & Garcia-Nunez, C. 1999 The effect of irrigation frequency on water and carbon relations in three cultivars of sweet pepper ( Capsicum chinense Jacq), in a tropical semiarid region Sci. Hort. 81 301 308
Karam, F. , Saliba, R. , Skaf, S. , Breidy, J. , Rouphael, Y. & Balendonck, J. 2011 Yield and water use of eggplants ( Solanum melongena L.) under full and deficit irrigation regimes Agr. Water Mgt. 98 1307 1316
Kirnak, H. , Tas, I. , Kaya, C. & Higgs, D. 2002 Effects of deficit irrigation on growth, yield, and fruit quality of eggplant under semi-arid conditions Austral. J. Agr. Res. 53 1367 1373
Larcher, W. 1995 Physiological plant ecology: Ecophysiological and stress physiology of functional groups. Springer, Berlin, Germany
Lei, S. , Yunzhou, Q. , Fengchao, J. , Changhai, S. , Chao, Y. , Yuxin, L. , Mengyu, L. & Baodi, D. 2009 Physiological mechanism contributing to efficient use of water in field tomato under different irrigation Plant Soil Environ. 55 128 133
Liu, Y.J. , Tong, Y.P. , Zhu, Y.G. , Ding, H. & Smith, E.A. 2006 Leaf chlorophyll readings as an indicator for spinach yield and nutritional quality with different nitrogen fertilizer applications J. Plant Nutr. 29 1207 1217
Locascio, S.J. , Olson, S.M. & Rhoads, F.M. 1989 Water quantity and time of N and K application for trickle-irrigated tomatoes J. Amer. Soc. Hort. Sci. 114 265 268
Madramootoo, C.A. & Rigby, M. 1991 Effects of trickle irrigation on the growth and sunscald of bell peppers ( Capsicum annuum L.) in southern Quebec Agr. Water Mgt. 19 181 189
Ngouajio, M. , Wang, G.Y. & Goldy, R. 2007 Withholding of drip irrigation between transplanting and flowering increases the yield of field-grown tomato under plastic mulch Agr. Water Mgt. 87 285 291
Ozores-Hampton, M. 2014 Conventional and specialty eggplant varieties in Florida. Horticultural Sciences Department, Univ. of Florida/Institute of Food and Agricultural Sciences. Document HS1243. 12 June 2015. < http://edis.ifas.ufl.edu/hs1243 >.
Pereira, L.S. , Allen, R.G. , Smith, M. & Raes, D. 2015 Crop evapotranspiration estimation with FAO56: Past and future Agr. Water Mgt. 147 4 20
Quintal Ortiz, W.C. , Perez-Gutierrez, A. , Latournerie Moreno, L. , May-Lara, C. , Ruiz Sanchez, E. & Martinez Chacon, A.J. 2012 Water use, water potential, and yield of habanero pepper ( Capsicum chinense Jacq.) Rev. Fitotec. Mex. 35 155 160
Sarker, B.C. , Hara, M. & Uemura, M. 2005 Proline synthesis, physiological responses and biomass yield of eggplants during and after repetitive soil moisture stress Sci. Hort. 103 387 402
Sezen, S.M. , Yazar, A. & Eker, S. 2006 Effect of drip irrigation regimes on yield and quality of field grown bell pepper Agr. Water Mgt. 81 115 131
Simonne, E.H. , Dukes, M.D. , Hochmuth, R.C. , Studstill, D.W. , Avezou, G. & Jarry, D. 2006 Scheduling drip irrigation for bell pepper grown with plasticulture J. Plant Nutr. 29 1729 1739
USDA 2013 United States standards for grades of eggplant. United States Department of Agriculture
Contributor Notes
Financial support was provided by the Georgia Agricultural Experiment Stations.
We thank John Silvoy, Jesús Bautista, and Nélida Bautista for their invaluable technical support. We also thank Peter Germishuizen from Lewis Taylor Farms, Ty Ty, GA, for donation of eggplant transplants. We appreciate the thorough review of the manuscript by Pat Conner, Tim Coolong, Erick Smith, and the anonymous reviewers.
Mention of trade names in this publication does not imply endorsement by the University of Georgia of products named, nor criticism of similar ones not mentioned. The cost of publishing this paper was defrayed in part by payment of page charges. Under postal regulations, this paper therefore must hereby be marked advertisement solely to indicate this fact.
1 Corresponding author. E-mail: [email protected] .
Headquarters:
1018 Duke Street
Alexandria, VA 22314
Phone : 703.836.4606
Email : [email protected]
© 2018-2024 American Society for Horticultural Science
- [66.249.64.20|195.158.225.244]
- 195.158.225.244
Character limit 500 /500
Health benefits and bioactive compounds of eggplant
Affiliations.
- 1 Izmir Institute of Technology, Department of Molecular Biology and Genetics, 35430 Urla Izmir, Turkey.
- 2 Mehmet Akif Ersoy University, Burdur Food Agriculture and Livestock Vocational School, 15030 Burdur, Turkey.
- 3 Izmir Institute of Technology, Department of Molecular Biology and Genetics, 35430 Urla Izmir, Turkey. Electronic address: [email protected].
- 4 Mount Holyoke College, Department of Biological Sciences, The Biochemistry Program, 50 College St, South Hadley, MA 01075, USA. Electronic address: [email protected].
- 5 Izmir Institute of Technology, Department of Molecular Biology and Genetics, 35430 Urla Izmir, Turkey. Electronic address: [email protected].
- PMID: 30064803
- DOI: 10.1016/j.foodchem.2018.06.093
Eggplant is a vegetable crop that is grown around the world and can provide significant nutritive benefits thanks to its abundance of vitamins, phenolics and antioxidants. In addition, eggplant has potential pharmaceutical uses that are just now becoming recognized. As compared to other crops in the Solanaceae, few studies have investigated eggplant's metabolic profile. Metabolomics and metabolic profiling are important platforms for assessing the chemical composition of plants and breeders are increasingly concerned about the nutritional and health benefits of crops. In this review, the historical background and classification of eggplant are shortly explained; then the beneficial phytochemicals, antioxidant activity and health effects of eggplant are discussed in detail.
Keywords: Bioactive compounds; Chemical composition; Eggplant; Metabolic profiling; Solanum melongena.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Publication types
- Antioxidants / isolation & purification
- Antioxidants / metabolism*
- Crops, Agricultural
- Phenols / isolation & purification
- Phenols / metabolism*
- Solanum melongena / chemistry*
- Antioxidants
Introduction: The Importance of Eggplant
- First Online: 31 May 2019
Cite this chapter

- Mark A. Chapman 3
Part of the book series: Compendium of Plant Genomes ((CPG))
782 Accesses
10 Citations
In this chapter, I highlight how the eggplant, whilst being globally dwarfed by other members of the Solanaceae, notably potato and tomato, offers a number of important ecological, evolutionary and agronomic features making it unique and interesting, warranting further study. It also highlights the parallels and differences between Solanaceous crops. The eggplant genome is in the process of being finalised, and once this is available to researchers, it is likely we will see a surge of papers utilising this resource for understanding the genetic basis of these important traits.
This is a preview of subscription content, log in via an institution to check access.
Access this chapter
- Available as PDF
- Read on any device
- Instant download
- Own it forever
- Available as EPUB and PDF
- Durable hardcover edition
- Dispatched in 3 to 5 business days
- Free shipping worldwide - see info
Tax calculation will be finalised at checkout
Purchases are for personal use only
Institutional subscriptions
Literature Cited
Bagnaresi P, Sala T, Irdani T, Scotto C, Lamontanara A, Beretta M, Rotino GL, Sestili S, Cattivelli L, Sabatini E (2013) Solanum torvum responses to the root-knot nematode Meloidogyne incognita . BMC Genom 14:540. https://doi.org/10.1186/1471-2164-14-540
Article CAS Google Scholar
Bahadur A, Rai N, Kumar R, Tiwari SK, Singh AK, Rai AK, Singh U, Patel PK, Tiwari V, Rai AB, Singh M, Singh BB (2015) Grafting tomato on eggplant as a potential tool to improve waterlogging tolerance in hybrid tomato. Veg Sci 42:82–87
Google Scholar
Bletsos F, Thanassoulopoulos C, Roupakias D (2003) Effect of grafting on growth, yield, and verticillium wilt of eggplant. HortScience 38:183–186
Article Google Scholar
Chapman MA (2015) Transcriptome sequencing and marker development for four underutilized legumes. Appl Plant Sci 3(2):1400111
Chapman MA, Burke JM (2006) Letting the gene out of the bottle: the population genetics of genetically modified crops. New Phytol 170(3):429–443
D’Arcy WG (1991) The solanaceae since 1976, with a review of its biogeography. In: Hawkes JG, Lester RN, Nee M, Estrada N (eds) Solanaceae III: taxonomy, chemistry, evolution. Royal Botanic Gardens, Kew, pp 75–137
Daunay MC (2008) Eggplant. In: Prohens J, Nuez F (eds) Vegetables II: fabaceae, liliaceae, solanaceae, and umbelliferae. Springer, New York, pp 163–220
Chapter Google Scholar
Daunay MC, Lester RN (1988) The usefulness of taxonomy for Solanaceae breeders, with special reference to the genus Solanum and to Solanum melongena L. (eggplant). Capsicum Newslett 7:70–79
Davidar P, Snow AA, Rajkumar M, Pasquet R, Daunay MC, Mutegi E (2015) The potential for crop to wild hybridization in eggplant ( Solanum melongena ; Solanaceae) in southern India. Am J Bot 102(1):129–139. https://doi.org/10.3732/ajb.1400404
Article PubMed Google Scholar
De Rosso VV, Morán Vieyra FE, Mercadante AZ, Borsarelli CD (2008) Singlet oxygen quenching by anthocyanin’s flavylium cations. Free Radical Res 42(10):885–891. https://doi.org/10.1080/10715760802506349
Dempewolf H, Baute G, Anderson J, Kilian B, Smith C, Guarino L (2017) Past and future use of wild relatives in crop breeding. Crop Sci 57(3):1070–1082. https://doi.org/10.2135/cropsci2016.10.0885
Doganlar S, Frary A, Daunay MC, Lester RN, Tanksley SD (2002) Conservation of gene function in the Solanaceae as revealed by comparative mapping of domestication traits in eggplant. Genetics 161(4):1713–1726
CAS PubMed PubMed Central Google Scholar
FAO (2017) FAOSTAT database collections ( http://faostat.fao.org ). Food and Agriculture Organization of the United Nations, Rome, Italy. Sept 2017
Fita A, Fioruci F, Plazas M, Rodriguez-Burruezo A, Prohens J (2015) Drought tolerance among accessions of eggplant and related species. Bull Univ Agric Sci Vet Med Cluj Napoca Hortic 72:461–462
Frodin DG (2004) History and concepts of big plant genera. Taxon 53:753–776
Gayoso C, Pomar F, Novo-Uzal E, Merino F, de Ilarduya OM (2010) The Ve-mediated resistance response of the tomato to Verticillium dahliae involves H 2 O 2 , peroxidase and lignins and drives PAL gene expression. BMC Plant Biol 10:232. https://doi.org/10.1186/1471-2229-10-232
Article CAS PubMed PubMed Central Google Scholar
Genova C, Schreinemachers P, Afari-Sefa V (2013) An impact assessment of AVRDC’s tomato grafting in Vietnam. AVRDC—The World Vegetable Center, Shanhua
Gousset C, Collonnier C, Mulya K, Mariska I, Rotino GL, Besse P, Servaes A, Sihachakr D (2005) Solanum torvum , as a useful source of resistance against bacterial and fungal diseases for improvement of eggplant ( S. melongena L.). Plant Sci 168(2):319–327. https://doi.org/10.1016/j.plantsci.2004.07.034
Gramazio P, Blanca J, Ziarsolo P, Herraiz FJ, Plazas M, Prohens J, Vilanova S (2016) Transcriptome analysis and molecular marker discovery in Solanum incanum and S. aethiopicum , two close relatives of the common eggplant ( Solanum melongena ) with interest for breeding. BMC Genom 17:300. https://doi.org/10.1186/s12864-016-2631-4
Gramazio P, Prohens J, Plazas M, Mangino G, Herraiz FJ, Garcia-Fortea E, Vilanova S (2018) Genomic tools for the enhancement of vegetable crops: a case in eggplant. Not Bot Horti Agrobo Cluj Napoca 46(1):1–13. https://doi.org/10.15835/nbha46110936
Groot AT, Dicke M (2002) Insect-resistant transgenic plants in a multi-trophic context. Plant J 31:387–406
Hautea DM, Taylo LD, Masanga APL, Sison MLJ, Narciso JO, Quilloy RB, Hautea RA, Shotkoski FA, Shelton AM (2016) Field performance of Bt eggplants ( Solanum melongena L.) in the Philippines: Cry1Ac expression and control of the eggplant fruit and shoot borer ( Leucinodes orbonalis Guenee). PLOS ONE 11(6):e0157498. https://doi.org/10.1371/journal.pone.0157498
Hawkes JG (1999) The economic importance of the family Solanaceae. In: Nee M, Symon DE, Lester RN, Jessop JP (eds) Solanaceae IV: advance in biology and utilization. Royal Botanic Gardens, Kew, pp 1–9
Herring RJ (2015) State science, risk and agricultural biotechnology: Bt cotton to Bt Brinjal in India. J Peasant Stud 42(1):159–186. https://doi.org/10.1080/03066150.2014.951835
Hirakawa H, Shirasawa K, Miyatake K, Nunome T, Negoro S, Ohyama A, Yamaguchi H, Sato S, Isobe S, Tabata S, Fukuoka H (2014) Draft genome sequence of eggplant ( Solanum melongena L.): the representative Solanum species indigenous to the Old World. DNA Res 21(6):649–660. https://doi.org/10.1093/dnares/dsu027
James C (2003) Global review of commercialized transgenic crops. Curr Sci 84(3):303–309
Kashyap A, Weber S (2013) Starch grain analysis and experiments provide insights into Harappan cooking practices. In: Abraham SA, Gullapalli P, Raczek TP, Rizvi UZ (eds) Connections and complexity: new approaches to the archaeology of South Asia. Left Coast Press, Taylor and Francis, Oxford, pp 177–193
Kaushik P, Prohens J, Vilanova S, Gramazio P, Plazas M (2016) Phenotyping of eggplant wild relatives and interspecific hybrids with conventional and phenomics descriptors provides insight for their potential utilization in breeding. Front Plant Sci 7:677. https://doi.org/10.3389/fpls.2016.00677
Article PubMed PubMed Central Google Scholar
Keatinge JDH, Lin LJ, Ebert AW, Chen WY, Hughes JdA, Luther GC, Wang JF, Ravishankar M (2014) Overcoming biotic and abiotic stresses in the Solanaceae through grafting: current status and future perspectives. Biol Agric Hortic 30(4):272–287. https://doi.org/10.1080/01448765.2014.964317
Klosterman SJ, Atallah ZK, Vallad GE, Subbarao KV (2009) Diversity, pathogenicity, and management of verticillium species. Annu Rev Phytopathol 47(1):39–62. https://doi.org/10.1146/annurev-phyto-080508-081748
Article CAS PubMed Google Scholar
Knapp S, Vorontsova MS, Prohens J (2013) Wild relatives of the eggplant ( Solanum melongena L.: Solanaceae): new understanding of species names in a complex group. PLoS ONE 8(2):e57039. https://doi.org/10.1371/journal.pone.0057039
Kouassi B, Prohens J, Gramazio P, Kouassi AB, Vilanova S, Galan-Avila A, Herraiz FJ, Kouassi A, Segui-Simarro JM, Plazas M (2016) Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant ( Solanum melongena ). Sci Hortic 213:199–207. https://doi.org/10.1016/j.scienta.2016.10.039
Li J, Ren L, Gao Z, Jiang MM, Liu Y, Zhou L, He YJ, Chen HY (2017) Combined transcriptomic and proteomic analysis constructs a new model for light-induced anthocyanin biosynthesis in eggplant ( Solanum melongena L.). Plant Cell Environ 40(12):3069–3087. https://doi.org/10.1111/pce.13074
Li J, He YJ, Zhou L, Liu Y, Jiang MM, Ren L, Chen HY (2018) Transcriptome profiling of genes related to light-induced anthocyanin biosynthesis in eggplant ( Solanum melongena L.) before purple color becomes evident. BMC Genom 19:324. https://doi.org/10.1186/s12864-018-4587-z
Lin Z, Li X, Shannon LM, Yeh C-T, Wang ML, Bai G, Peng Z, Li J, Trick HN, Clemente TE, Doebley J, Schnable PS, Tuinstra MR, Tesso TT, White F, Yu J (2012) Parallel domestication of the Shattering1 genes in cereals. Nat Genet 44(6):720–724. https://doi.org/10.1038/ng.2281
Liu B, Zeng Q, Yan FM, Xu HG, Xu CR (2005) Effects of transgenic plants on soil microorganisms. Plant Soil 271(1–2):1–13
Liu J, Zheng Z, Zhou X, Feng C, Zhuang Y (2015) Improving the resistance of eggplant ( Solanum melongena ) to Verticillium wilt using wild species Solanum linnaeanum . Euphytica 201(3):463–469. https://doi.org/10.1007/s10681-014-1234-x
Lotito SB, Frei B (2006) Consumption of flavonoid-rich foods and increased plasma antioxidant capacity in humans: Cause, consequence, or epiphenomenon? Free Radical Biol Med 41(12):1727–1746. https://doi.org/10.1016/j.freeradbiomed.2006.04.033
Ma C, Dastmalchi K, Whitaker BD, Kennelly EJ (2011) Two new antioxidant malonated caffeoylquinic acid isomers in fruits of wild eggplant relatives. J Agric Food Chem 59(17):9645–9651. https://doi.org/10.1021/jf202028y
Meyer RS, DuVal AE, Jensen HR (2012a) Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol 196(1):29–48. https://doi.org/10.1111/j.1469-8137.2012.04253.x
Meyer RS, Karol KG, Little DP, Nee MH, Litt A (2012b) Phylogeographic relationships among Asian eggplants and new perspectives on eggplant domestication. Mol Phylogen Evol 63:685–701
Meyer RS, Bamshad M, Fuller DQ, Litt A (2014) Comparing medicinal uses of eggplant and related solanaceae in china, india, and the Philippines suggests the independent development of uses, cultural diffusion, and recent species substitutions. Econ Bot 68(2):137–152. https://doi.org/10.1007/s12231-014-9267-6
Meyer RS, Whitaker BD, Little DP, Wu S-B, Kennelly EJ, Long C-L, Litt A (2015) Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 115:194–206. https://doi.org/10.1016/j.phytochem.2015.02.006
Morrell PL, Buckler ES, Ross-Ibarra J (2011) Crop genomics: advances and applications. Nat Rev Genet 13:85. https://doi.org/10.1038/nrg3097
Navasero MV, Candano RN, Hautea DM, Hautea RA, Shotkoski FA, Shelton AM (2016) Assessing potential impact of Bt eggplants on non-target arthropods in the Philippines. PLoS ONE 11(10):e0165190. https://doi.org/10.1371/journal.pone.0165190
Ozsolak F, Milos PM (2010) RNA sequencing: advances, challenges and opportunities. Nat Rev Genet 12:87. https://doi.org/10.1038/nrg2934
Piquerez SJM, Harvey SE, Beynon JL, Ntoukakis V (2014) Improving crop disease resistance: lessons from research on Arabidopsis and tomato. Front Plant Sci 5:671. https://doi.org/10.3389/fpls.2014.00671
Qtu J, Hallmann J, Kokalis-Burelle N, Weaver DB, Rodriguez-Kabana R, Tuzun S (1997) Activity and differential induction of chitinase isozymes in soybean cultivars resistant or susceptible to root-knot nematodes. J Nematol 29(4):523–530
Rai NP, Rai GK, Kumar S, Kumari N, Singh M (2013) Shoot and fruit borer resistant transgenic eggplant ( Solanum melongena L.) expressing cry1Aa3 gene: development and bioassay. Crop Protect 53:37–45. https://doi.org/10.1016/j.cropro.2013.06.005
Ranil RHG, Prohens J, Aubriot X, Niran HML, Plazas M, Fonseka RM, Vilanova S, Fonseka HH, Gramazio P, Knapp S (2017) Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant ( S. melongena L.): a review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet Resour Crop Evol 64(7):1707–1722. https://doi.org/10.1007/s10722-016-0467-z
Salgon S, Jourda C, Sauvage C, Daunay M-C, Reynaud B, Wicker E, Dintinger J (2017) Eggplant resistance to the Ralstonia solanacearum species complex involves both broad-spectrum and strain-specific quantitative trait loci. Front Plant Sci 8:828
Sato S, Tabata S, Hirakawa H, Asamizu E, Shirasawa K, Isobe S, Kaneko T, Nakamura Y, Shibata D, Aoki K, Egholm M, Knight J, Bogden R, Li C, Shuang Y, Xu X, Pan S, Cheng S, Liu X, Ren Y, Wang J, Albiero A, Dal Pero F, Todesco S, Van Eck J, Buels RM, Bombarely A, Gosselin JR, Huang M, Leto JA, Menda N, Strickler S, Mao L, Gao S, Tecle IY, York T, Zheng Y, Vrebalov JT, Lee J, Zhong S, Mueller LA, Stiekema WJ, Ribeca P, Alioto T, Yang W, Huang S, Du Y, Zhang Z, Gao J, Guo Y, Wang X, Li Y, He J, Li C, Cheng Z, Zuo J, Ren J, Zhao J, Yan L, Jiang H, Wang B, Li H, Li Z, Fu F, Chen B, Han B, Feng Q, Fan D, Wang Y, Ling H, Xue Y, Ware D, McCombie WR, Lippman ZB, Chia J-M, Jiang K, Pasternak S, Gelley L, Kramer M, Anderson LK, Chang S-B, Royer SM, Shearer LA, Stack SM, Rose JKC, Xu Y, Eannetta N, Matas AJ, McQuinn R, Tanksley SD, Camara F, Guigo R, Rombauts S, Fawcett J, Van de Peer Y, Zamir D, Liang C, Spannagl M, Gundlach H, Bruggmann R, Mayer K, Jia Z, Zhang J, Ye Z, Bishop GJ, Butcher S, Lopez-Cobollo R, Buchan D, Filippis I, Abbott J, Dixit R, Singh M, Singh A, Pal JK, Pandit A, Singh PK, Mahato AK, Dogra V, Gaikwad K, Sharma TR, Mohapatra T, Singh NK, Causse M, Rothan C, Schiex T, Noirot C, Bellec A, Klopp C, Delalande C, Berges H, Mariette J, Frasse P, Vautrin S, Zouine M, Latche A, Rousseau C, Regad F, Pech J-C, Philippot M, Bouzayen M, Pericard P, Osorio S, Fernandez del Carmen A, Monforte A, Granell A, Fernandez-Munoz R, Conte M, Lichtenstein G, Carrari F, De Bellis G, Fuligni F, Peano C, Grandillo S, Termolino P, Pietrella M, Fantini E, Falcone G, Fiore A, Giuliano G, Lopez L, Facella P, Perrotta G, Daddiego L, Bryan G, Orozco M, Pastor X, Torrents D, van Schriek KNVMGM, Feron RMC, van Oeveren J, de Heer P, daPonte L, Jacobs-Oomen S, Cariaso M, Prins M, van Eijk MJT, Janssen A, van Haaren MJJ, Jo S-H, Kim J, Kwon S-Y, Kim S, Koo D-H, Lee S, Hur C-G, Clouser C, Rico A, Hallab A, Gebhardt C, Klee K, Joecker A, Warfsmann J, Goebel U, Kawamura S, Yano K, Sherman JD, Fukuoka H, Negoro S, Bhutty S, Chowdhury P, Chattopadhyay D, Datema E, Smit S, Schijlen EWM, van de Belt J, van Haarst JC, Peters SA, van Staveren MJ, Henkens MHC, Mooyman PJW, Hesselink T, van Ham RCHJ, Jiang G, Droege M, Choi D, Kang B-C, Kim BD, Park M, Kim S, Yeom S-I, Lee Y-H, Choi Y-D, Li G, Gao J, Liu Y, Huang S, Fernandez-Pedrosa V, Collado C, Zuniga S, Wang G, Cade R, Dietrich RA, Rogers J, Knapp S, Fei Z, White RA, Thannhauser TW, Giovannoni JJ, Angel Botella M, Gilbert L, Gonzalez R, Goicoechea JL, Yu Y, Kudrna D, Collura K, Wissotski M, Wing R, Schoof H, Meyers BC, Gurazada AB, Green PJ, Mathur S, Vyas S, Solanke AU, Kumar R, Gupta V, Sharma AK, Khurana P, Khurana JP, Tyagi AK, Dalmay T, Mohorianu I, Walts B, Chamala S, Barbazuk WB, Li J, Guo H, Lee T-H, Wang Y, Zhang D, Paterson AH, Wang X, Tang H, Barone A, Chiusano ML, Ercolano MR, D’Agostino N, Di Filippo M, Traini A, Sanseverino W, Frusciante L, Seymour GB, Elharam M, Fu Y, Hua A, Kenton S, Lewis J, Lin S, Najar F, Lai H, Qin B, Qu C, Shi R, White D, White J, Xing Y, Yang K, Yi J, Yao Z, Zhou L, Roe BA, Vezzi A, D’Angelo M, Zimbello R, Schiavon R, Caniato E, Rigobello C, Campagna D, Vitulo N, Valle G, Nelson DR, De Paoli E, Szinay D, de Jong HH, Bai Y, Visser RGF, Lankhorst RMK, Beasley H, McLaren K, Nicholson C, Riddle C, Gianese G, Tomato Genome Consortium (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485(7400):635–641. https://doi.org/10.1038/nature11119
Singh AK, Rai GK, Singh M, Dubey SK (2013) Bacterial community structure in the rhizosphere of a Cry1Ac Bt-Brinjal crop and comparison to its non-transgenic counterpart in the tropical soil. Microb Ecol 66(4):927–939. https://doi.org/10.1007/s00248-013-0287-z
Stommel JR, Whitaker BD (2003) Phenolic acid composition of eggplant fruit in a germplasm core subset. J Am Soc Hort Sci 128:704–710
Toppino L, Valè G, Rotino GL (2008) Inheritance of Fusarium wilt resistance introgressed from Solanum aethiopicum Gilo and Aculeatum groups into cultivated eggplant ( S. melongena ) and development of associated PCR-based markers. Mol Breed 22(2):237–250. https://doi.org/10.1007/s11032-008-9170-x
Wang JX, Gao TG, Knapp S (2008) Ancient Chinese literature reveals pathways of eggplant domestication. Ann Bot 102(6):891–897. https://doi.org/10.1093/aob/mcn179
Weese TL, Bohs L (2010) Eggplant origins: out of Africa, into the Orient. Taxon 59:49–56
White OW, Doo B, Carine MA, Chapman MA (2016) Transcriptome sequencing and simple sequence repeat marker development for three Macaronesian endemic plant species. Appl Plant Sci 4(8):1600050. https://doi.org/10.3732/apps.1600050
Wilusz JE, Sunwoo H, Spector DL (2009) Long noncoding RNAs: functional surprises from the RNA world. Genes Dev 23(13):1494–1504. https://doi.org/10.1101/gad.1800909
Xu L, Zhu L, Tu L, Liu L, Yuan D, Jin L, Long L, Zhang X (2011a) Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-seq-dependent transcriptional analysis and histochemistry. J Exp Bot 62(15):5607–5621. https://doi.org/10.1093/jxb/err245
Xu X, Pan S, Cheng S, Zhang B, Mu D, Ni P, Zhang G, Yang S, Li R, Wang J, Orjeda G, Guzman F, Torres M, Lozano R, Ponce O, Martinez D, De la Cruz G, Chakrabarti SK, Patil VU, Skryabin KG, Kuznetsov BB, Ravin NV, Kolganova TV, Beletsky AV, Mardanov AV, Di Genova A, Bolser DM, Martin DMA, Li G, Yang Y, Kuang H, Hu Q, Xiong X, Bishop GJ, Sagredo B, Mejia N, Zagorski W, Gromadka R, Gawor J, Szczesny P, Huang S, Zhang Z, Liang C, He J, Li Y, He Y, Xu J, Zhang Y, Xie B, Du Y, Qu D, Bonierbale M, Ghislain M, del Rosario Herrera M, Giuliano G, Pietrella M, Perrotta G, Facella P, O’Brien K, Feingold SE, Barreiro LE, Massa GA, Diambra L, Whitty BR, Vaillancourt B, Lin H, Massa A, Geoffroy M, Lundback S, DellaPenna D, Buell CR, Sharma SK, Marshall DF, Waugh R, Bryan GJ, Destefanis M, Nagy I, Milbourne D, Thomson SJ, Fiers M, Jacobs JME, Nielsen KL, Sonderkaer M, Iovene M, Torres GA, Jiang J, Veilleux RE, Bachem CWB, de Boer J, Borm T, Kloosterman B, van Eck H, Datema E, Hekkert BTL, Goverse A, van Ham RCHJ, Visser RGF, Potato Genome Sequencing Consortium (2011b) Genome sequence and analysis of the tuber crop potato. Nature 475(7355):189–194. https://doi.org/10.1038/nature10158
Yang X, Cheng Y-F, Deng C, Ma Y, Wang Z-W, Chen X-H, Xue L-B (2014) Comparative transcriptome analysis of eggplant ( Solanum melongena L.) and turkey berry ( Solanum torvum Sw.): phylogenomics and disease resistance analysis. BMC Genom 15(1):412. https://doi.org/10.1186/1471-2164-15-412
Zhang YJ, Chu GH, Hu ZL, Gao Q, Cui BL, Tian SB, Wang B, Chen GP (2016) Genetically engineered anthocyanin pathway for high health-promoting pigment production in eggplant. Mol Breed 36(5):54. https://doi.org/10.1007/s11032-016-0454-2
Zhou XH, Bao SY, Liu J, Zhuang Y (2016) De novo sequencing and analysis of the transcriptome of the wild eggplant species Solanum aculeatissimum in response to verticillium dahliae. Plant Mol Biol Rep 34(6):1193–1203. https://doi.org/10.1007/s11105-016-0998-7
Download references
Author information
Authors and affiliations.
Biological Sciences, University of Southampton, Southampton, SO17 1BJ, UK
Mark A. Chapman
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Mark A. Chapman .
Editor information
Editors and affiliations.
Biological Sciences, University of Southampton, Southampton, UK
Rights and permissions
Reprints and permissions
Copyright information
© 2019 Springer Nature Switzerland AG
About this chapter
Chapman, M.A. (2019). Introduction: The Importance of Eggplant. In: Chapman, M. (eds) The Eggplant Genome. Compendium of Plant Genomes. Springer, Cham. https://doi.org/10.1007/978-3-319-99208-2_1
Download citation
DOI : https://doi.org/10.1007/978-3-319-99208-2_1
Published : 31 May 2019
Publisher Name : Springer, Cham
Print ISBN : 978-3-319-99207-5
Online ISBN : 978-3-319-99208-2
eBook Packages : Biomedical and Life Sciences Biomedical and Life Sciences (R0)
Share this chapter
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Publish with us
Policies and ethics
- Find a journal
- Track your research
REVIEW article
World vegetable center eggplant collection: origin, composition, seed dissemination and utilization in breeding.

- 1 World Vegetable Center, Tainan, Taiwan
- 2 Vegetable Crops Research Department, Agriculture Research Center, Horticulture Research Institute, Giza, Egypt
- 3 Faculty of Applied Ecology and Agricultural Sciences, Inland Norway University of Applied Sciences, Elverum, Norway
- 4 Instituto de Conservación y Mejora de la Agrodiversidad Valenciana, Universitat Politècnica de València, Valencia, Spain
- 5 Horticulture Department, Faculty of Agriculture, University of Kafrelsheikh, Kafr El-Sheikh, Egypt
Eggplant is the fifth most economically important solanaceous crop after potato, tomato, pepper, and tobacco. Apart from the well-known brinjal eggplant ( Solanum melongena L.), two other under-utilized eggplant species, the scarlet eggplant ( S. aethiopicum L.) and the gboma eggplant ( S. macrocarpon L.) are also cultivated. The taxonomy and identification of eggplant wild relatives is challenging for breeders due to the large number of related species, but recent phenotypic and genetic data and classification in primary, secondary, and tertiary genepools, as well as information on the domestication process and wild progenitors, facilitates their utilization in breeding. The World Vegetable Center (WorldVeg) holds a large public germplasm collection of eggplant, which includes the three cultivated species and more than 30 eggplant wild relatives, with more than 3,200 accessions collected from 90 countries. Over the last 15 years, more than 10,000 seed samples from the Center's eggplant collection have been shared with public and private sector entities, including other genebanks. An analysis of the global occurrences and genebank holdings of cultivated eggplants and their wild relatives reveals that the WorldVeg genebank holds the world's largest public collection of the three cultivated eggplant species. The composition, seed dissemination and utilization of germplasm from the Center's collection are highlighted. In recent years more than 1,300 accessions of eggplant have been characterized for yield and fruit quality parameters. Further screening for biotic and abiotic stresses in eggplant wild relatives is a priority, as is the need to amass more comprehensive knowledge regarding wild relatives' potential for use in breeding. However, as is the case for many other crops, wild relatives are highly under-represented in the global conservation system of eggplant genetic resources.
Introduction
Brinjal eggplant ( Solanum melongena L.) is a warm-weather crop mostly cultivated in tropical and subtropical regions of the world. Two other cultivated eggplant species, the scarlet eggplant ( S. aethiopicum L.) and the gboma eggplants ( S. macrocarpon L.), are less known but have local importance in sub-Saharan Africa ( Schippers, 2000 ; Daunay and Hazra, 2012 ). Based on data from 2014, the global production of eggplant is around 50 million tons annually, with a net value of more than US$10 billion a year, which makes it the fifth most economically important solanaceous crop after potato, tomato, pepper, and tobacco ( FAO, 2014 ). The top five producing countries are China (28.4 million tons; 57% of world's total), India (13.4 million tons; 27% of world's total), Egypt (1.2 million tons), Turkey (0.82 million tons), and Iran (0.75 million tons). In Asia and the Mediterranean, eggplant ranks among the top five most important vegetable crops ( Frary et al., 2007 ).
Regarding nutritional value, eggplant has a very low caloric value and is considered among the healthiest vegetables for its high content of vitamins, minerals and bioactive compounds for human health ( Raigón et al., 2008 ; Plazas et al., 2014b ; Docimo et al., 2016 ). In this respect, eggplant is ranked among the top 10 vegetables in terms of oxygen radical absorbance capacity ( Cao et al., 1996 ). The bioactive properties of eggplant are mostly associated with high content in phenolic compounds ( Plazas et al., 2013 ), which are mostly phenolic acids, particularly chlorogenic acid in the fruit flesh ( Stommel et al., 2015 ) and anthocyanins in the fruit skin ( Mennella et al., 2012 ). Both phenolic acids and anthocyanins have multiple properties beneficial for human health ( Plazas et al., 2013 ; Braga et al., 2016 ).
Farmers need improved eggplant varieties for sustainable production and adaptation to climate change challenges. Because eggplant has a relatively long growth period, it is more exposed than other vegetable crops to a broad range of plant diseases, pests, nematodes, and weeds. The most common diseases include bacterial wilt, verticillium wilt, fusarium wilt, anthracnose fruit rot, alternaria rot, damping off, Phytophthora blight, phomopsis blight and fruit rot, leaf spot, little leaf of brinjal, and mosaic ( Rotino et al., 1997 ). Eggplant is also subject to attack by numerous insect pests including mites, whiteflies, aphids, eggplant fruit, and shoot borer, leafhopper, thrips, spotted beetles, leaf roller, stem borer, and blister beetle ( Rotino et al., 1997 ; Medakker and Vijayaraghavan, 2007 ). Unpredictable weather with extreme temperatures, drought or flooding can reduce yield and fruit quality. In general, eggplant breeding programs aim to develop high-yielding varieties, mostly F 1 hybrids, with high fruit quality, shelf-life and resistance to major disease and insect pests, and with broad adaptation to environmental stress ( Daunay and Hazra, 2012 ).
Access to genetic diversity is fundamental for any breeding program. In this paper, we review the diversity and genetic resources of eggplant. As a point of departure, we examine the taxonomy and relationships of the crop and its wild relatives, as well as the domestication of cultivated eggplant. The relationships among wild, semi-domesticated, and cultivated eggplant are intricate, and the origin, evolution, and migration are incompletely understood ( Levin et al., 2006 ; Meyer et al., 2012 ). Here, we limit ourselves to identify global occurrences and regions of diversity. A key section is the overview of global genebank holdings of cultivated eggplant and their wild relatives. As we shall demonstrate, for such plants the collection at the WorldVeg is of paramount importance. Composition, seed dissemination and utilization of germplasm from this collection are presented and discussed. The importance of safeguarding and evaluating wild relatives is highlighted, as crop wild relatives are highly under-represented in the global conservation system of plant genetic resources and may harbor important genes for resistance or tolerance to biotic and abiotic stresses.
Taxonomy, Wild Relatives, and Domestication of Eggplant
Eggplants are berry-producing vegetables belonging to the large Solanaceae family (nightshade family), which contains ~3,000 species distributed in some 90 genera ( Vorontsova and Knapp, 2012 ). Out of these Solanum L. is the largest one, with around 1,500 species ( Frodin, 2004 ) including globally important crops such as potato ( Solanum tuberosum L.) and tomato ( Solanum lycopersicum L.), as well as many other minor crops. Most taxa of Solanum genus have a basic chromosome number of n = 12 ( Chiarini et al., 2010 ).
The Solanum genus is mega-diverse and can be divided into 13 clades, where eggplant is the member of the large and taxonomically challenging Leptostemonum clade (subgenus Leptostemonum Bitter; Knapp et al., 2013 ), which is commonly known as the “spiny Solanum ” group due to the presence of sharp epidermal prickles on stems and leaves ( Vorontsova et al., 2013 ). The subgenus Leptostemonum contains around 450 currently recognized species distributed worldwide ( Knapp et al., 2013 ), many of which originated in the New World ( Vorontsova and Knapp, 2012 ). All three cultivated eggplant species have the Old World in origin (Figure 1 ). The Old World (Africa and Eurasia) and Australia, are home to more than 300 Solanum species ( Levin et al., 2006 ; Vorontsova and Knapp, 2016 ). Solanum melongena and S. macrocarpon are usually included in section Melongena Dunal ( Lester and Daunay, 2003 ; Lester et al., 2011 ), whereas S. aethiopicum is assigned to section Oliganthes (Dunal) Bitter ( Lester, 1986 ).

Figure 1 . Schematic representation of taxonomic relationships between the cultivated brinjal eggplant ( Solanum melongena ) and other cultivated (scarlet eggplant, S. aethiopicum ; and gboma eggplant, S. macrocarpon ) and wild relatives from the genus Solanum based on Nee (1999) , Levin et al. (2006) , Weese and Bohs (2010) , Stern et al. (2011) , Knapp et al. (2013) , Syfert et al. (2016) , and Vorontsova and Knapp (2016) . For each of the species and groups it is indicated if they are part of the primary (GP1), secondary (GP2), or tertiary (GP3) brinjal eggplant genepools. The three cultivated species are indicated with an asterisk.
Solanum melongena is characterized by large morphological diversity, and frequently it has been considered as the same taxonomic species than its wild ancestor S. insanum L. ( Ranil et al., 2017 ). Four taxonomically informal groups, labeled E–H, were considered by Lester and Hasan (1991) to describe the different types of wild and weedy eggplant as well as their distribution (Table 1 ). However, these four groups are presently considered as representing two different species: the cultivated eggplant S. melongena and its wild ancestor S. insanum ( Knapp et al., 2013 ). In this way, groups E and F corresponding to extremely prickly and plants that grow wild or weedy in India and Southeast Asia are now included within S. insanum ( Ranil et al., 2017 ). The plants of group G correspond to primitive eggplant cultivars, with small fruits, while the plants of group H are less prickly than other groups and consist of large-fruited landraces and modern cultivars ( Daunay et al., 2001 ; Weese and Bohs, 2010 ; Table 1 ). Both groups, G and H, constitute S. melongena ( Knapp et al., 2013 ). Some studies ( Hurtado et al., 2012 ; Vilanova et al., 2012 ; Cericola et al., 2013 ) have also pointed to a genetic and morphological differentiation between Occidental (eggplants from the Mediterranean area, North of Africa, and Middle East) and Oriental (from southeast and eastern Asia).

Table 1 . Cultivated eggplants (brinjal eggplant, S. melongena L.; scarlet eggplant, S. anguivi L.; gboma eggplant, S. macrocarpon L.) and their wild relatives from the primary genepool, which correspond to their wild ancestors ( S. insanum L. for brinjal eggplant, S. anguivi for scarlet eggplant, and S. dasyphyllum for gboma eggplant) ( Lester, 1986 ; Lester and Niakan, 1986 ; Bukenya and Carasco, 1994 ; Schippers, 2000 ; Daunay et al., 2001 ; Weese and Bohs, 2010 ; Meyer et al., 2012 ; Knapp et al., 2013 ; Vorontsova and Knapp, 2016 ).
Solanum aethiopicum is also hyper-variable and is classified into four cultivar groups (Gilo, Shum, Kumba, and Aculeatum; Table 1 ) based on morphological characteristics and use ( Lester, 1986 ). The Gilo group has edible fruits with different shapes, color, and size, and hairy, inedible leaves; the Shum group has glabrous and small leaves that are eaten as a green vegetable but the fruits are inedible; the Kumba group has glabrous leaves and flattened large fruits, which are edible; the Aculeatum group, on the other hand, has more prickliness than other groups with flat-shaped fruit, and are used as ornamentals ( Lester, 1986 ; Prohens et al., 2012 ; Plazas et al., 2014a ).
Solanum macrocarpon is cultivated both for its leaves and fruits ( Schippers, 2000 ; Maundu et al., 2009 ; Table 1 ). The species is less morphologically diverse than S. melongena and S. aethiopicum ( Plazas et al., 2014a ).
Although, recent information exists on domestication of eggplants, there are still many unanswered questions about this process. Vavilov (1951) considered S. melongena as being native to the “Indo-Chinese center of origin.” However, recent evidence suggests that brinjal eggplant had a multiple independent domestication from S. insanum , which is naturally distributed in tropical Asia from Madagascar to the Philippines ( Knapp et al., 2013 ) in several centers of domestication ( Meyer et al., 2012 ). Although, the evidence of cultivation of eggplant in both India and China is equally old, archeological evidence suggests that utilization of wild eggplants may have started earlier in India than China, with a subsequent additional and independent center of domestication in the Philippines ( Meyer et al., 2012 ). Around the eighth century, eggplant spread eastward to Japan and then westward along the Silk Road into Western Asia, Europe, and Africa by Arab traders during the fourteenth century, then it was introduced into America soon after Europeans arrived there ( Prohens et al., 2005 ) and later expanded into other parts of world. Much less is known on the domestication of the scarlet and gboma eggplants. Both species were domesticated in Africa, from its respective wild ancestors, which are S. anguivi Lam. in the case of S. aethiopicum ( Lester and Niakan, 1986 ) and S. dasyphyllum Schumach. and Thonn. in the case of S. macrocarpon ( Bukenya and Carasco, 1994 ). Hybrids between cultivated eggplants and their respective wild ancestors are fully fertile ( Lester and Thitai, 1989 ; Bukenya and Carasco, 1994 ; Plazas et al., 2016 ).
Solanum melongena and the two other cultivated eggplants are related to a large number of wild species ( Vorontsova et al., 2013 ; Syfert et al., 2016 ) that may serve as sources of variation for breeding programs, in particular for traits related to adaptation to climate change but also pest and disease resistance ( Rotino et al., 2014 ). Some of these species are listed in Table 2 . Although, the brinjal eggplant is considered to be a vegetable of Asian origin, most eggplant wild relatives are from Africa ( Weese and Bohs, 2010 ). Wild eggplants produce small, bitter, multi-seeded fruits, almost always inedible, and the plant is generally very spiny. Some of them possess high levels of chlorogenic acid and other bioactive compounds, which may have potential interest for human health ( Meyer et al., 2015 ). The wild relatives of eggplant are one of the most variable and intricate groups, in regards to their taxonomic and phylogenetic relationships ( Vorontsova et al., 2013 ). Based on crossing and biosystematics data, nine wild species, together with S. melongena , form the “eggplant complex,” which includes the cultivated brinjal eggplant and its closest eggplant wild relatives ( Knapp et al., 2013 ). Wild relatives can be classified based on their crossability with cultivated species (genepool concept) into primary, secondary, and tertiary genepools ( Harlan and de Wet, 1971 ). The primary genepool (GP1) of brinjal eggplant consists of cultivated eggplant and its wild ancestor S. insanum ( Ranil et al., 2017 ) which can be crossed easily and produce normal fertile hybrids ( Plazas et al., 2016 ). The secondary genepool (GP2) includes a large number (over 40) wild relatives that can be crossed or are phylogenetically close to brinjal eggplant, but the success of the crosses and the viability or fertility of the hybrids with the brinjal eggplant may be reduced. For example, some interspecific hybrids derived from GP2 are partly sterile or weak due to reproductive barriers such as S. dasyphyllum, S. linnaeanum Hepper & P.-M. L. Jaeger or S. tomentosum L. ( Rotino et al., 2014 ; Kouassi et al., 2016 ). The tertiary genepool (GP3) includes more distantly related species, including New World species, which are used in breeding programs for their resistance features, but crossing needs specific breeding techniques to succeed (e.g., S. torvum Sw., S. elaeagnifolium Cav., and S. sisymbriifolium Lam.; Kouassi et al., 2016 ; Plazas et al., 2016 ; Syfert et al., 2016 ).

Table 2 . Cultivated eggplant and wild relatives, number of occurrences, their regions and number of conserved accessions globally and at the World Vegetable Center (WorldVeg).
Global Occurrences and Genebank Conservation of Eggplant and Wild Relatives
In the following section we review the current status of eggplant genetic resources including the cultivated species and their most recognized wild relatives using information collected from biodiversity, herbarium, and genebank databases. The Global Biodiversity Information Facility (GBIF) was applied to review the number of recorded occurrences, which can be natural populations, herbarium samples, or other biodiversity records ( GBIF, 2017 ). Scientific names were used as a filter in the search function. The total numbers of records per species were noted, as were clusters of occurrences that were identified visually by applying the database map function. The main cluster of S. melongena was in India, with more than 5,000 of the total number of around 18,000 occurrences. Other clusters were in Turkey, Southeast Asia, and Spain, while the main cluster of occurrences of S. aethiopicum and S. macrocarpon was in West Africa, with a total of 1,288 and 443 occurrences, respectively. Based on the literature of previous studies and characterization data available at the WorldVeg, a list of 35 crop wild relatives was included in this review, which had ~100 ( S. repandum G. Forst.) to more than 7,000 occurrences ( S. torvum ) on a global scale recorded by GBIF (Table 2 ). Important regions for wild relatives vary depending on the species, but include all continents; Latin America, Asia, and Africa are the most common areas for wild relatives.
The Global Gateway to Genetic Resources ( GENESYS, 2017 ) was applied to review the number of conserved genebank accessions. The database includes more than 3 million accessions, which is less than half of the estimated number of more than 7 million accessions that are conserved globally ( FAO, 2010 ). Although, not all national genebanks report to Genesys, we still used the information for reviewing global holdings. Scientific names were used as a filter in the search function of the database, and the most important holding institutions were identified from the summary function of the database. Additional sources were reviewed to try to capture important collections outside Genesys, including national genebank databases and the database for Svalbard Global Seed Vault ( SGSV, 2017 ). The WorldVeg plays a major role in the conservation and distribution of vegetable germplasm, holding 60,387 accessions comprising 173 genera and 440 species from 151 countries of origin ( AVGRIS, 2017 ).
In total, 5,665 accessions of S. melongena , 798 accessions of S. aethiopicum and 169 accessions of S. macrocarpon were reported by GENESYS (2017) . Important national eggplant collections not reporting to GENESYS are at the National Bureau of Plant Genetic Resources in India and the Institute of Vegetables and Flowers in China. Data from such collections were not included in our study. The largest collections of these three cultivated species were those of the WorldVeg [2,212 accessions of S. melongena (39%), 481 accessions of S. aethiopicum (60%), and 63 accessions of S. macrocarpon (37%)], followed by the Plant Genetic Resources Conservation Unit at the University of Georgia, USDA-ARS (close to 800 accessions of S. melongena ) and the Centre for Genetic Resources at the Netherlands Plant Research International (373 accessions of S. melongena ; GENESYS, 2017 ). The N. I. Vavilov Research Institute of Plant Genetic Resource in Russia has a significant eggplant collection with more than 500 S. melongena accessions. The conservation of wild species ranged from a few accessions (e.g., S. rigescentoides Hutch.) to 167 accessions ( S. incanum L.). None of the wild species had large collections. Interestingly, the WorldVeg has the largest collections for S. aculeatissimum Jacq. (46 accessions, 71%), S. anguivi (28 accessions, 23%), S. capense L. (3 accessions, 38%), S. ferox L. (11 accessions, 38%), S. indicum L. (12 accessions, 92%), S. insanum (11 accessions, 100%), S. lasiocarpum Dunal (31 accessions, 74%), S. stramoniifolium Jacq. (10 accessions, 63%), S. torvum (112 accessions, 85%), S. trilobatum L. (10 accessions, 71%), S. viarum Dunal (16 accessions, 27%), S. violaceum Ortega (49 accessions, 77%), and S. xanthocarpum Schrad. & J. C. Wendl. (18 accessions, 90%) ( GENESYS, 2017 ). The low number of accessions identified as S. insanum in the collections is surprising, taking into account that it is quite abundant and the progenitor of eggplant ( Knapp et al., 2013 ; Ranil et al., 2017 ). This is probably caused by the fact that many S. insanum accessions are probably conserved as S. melongena , as both species have often been considered as being a single species ( S. melongena ; e.g., Lester and Hasan, 1991 ). Also, the correct classification of accessions under “ S. indicum L.” should be determined, as this name was rejected in 1978 as it was used to refer to two clearly distinct species, the African S. anguivi and the Asian S. violaceum ( Vorontsova and Knapp, 2016 ).
According to our analysis, wild eggplants are greatly under-represented in ex situ repositories. Such findings are also reported by Castañeda-Álvarez et al. (2016) , where eggplants were among the crops whose wild genepools are highly under-represented. Indeed, there is a need for conducting collection missions and conservation actions for eggplant wild relatives (Conservation gaps, http://www.cwrdiversity.org/conservation-gaps/ , Accessed February 30, 2017).
Eggplant Germplasm Dissemination from the World Vegetable Center
As demonstrated in the previous section, the collection at the WorldVeg is the most significant eggplant collection worldwide. Eggplant is the Center's third most widely distributed vegetable crop after pepper and tomato. A total of 11,383 germplasm samples were distributed from WorldVeg headquarters to 90 countries from the period 2000 to 2017. Most of these were of S. melongena (10,519 samples; 92.4%), followed by S. aethiopicum (738 samples; 6.4%) and S. macrocarpon (126 samples; 2.2%; Table 3 ). These accessions correspond to landraces and traditional cultivars with significant diversity in plant morphology, fruit types and colors, and resistance to biotic and abiotic stresses. The largest share of germplasm samples went to other genebanks (7,042 samples; 61.8%), followed by National Agricultural Research & Extension System/Government (NARES) (2,154 samples; 18.9%), internal distribution to WorldVeg scientists (703 samples; 6.1%), and seed companies (503 samples; 4.4%).

Table 3 . The World Vegetable Center seed distribution of cultivated eggplant by recipient category during the period 2000–2017.
The large morphological diversity of the WorldVeg collection is matched by the identification of traits of significant agronomic interest. WorldVeg has compiled and maintained the world's largest germplasm collection of eggplant, and national genebanks and institutions from around the globe have requested and received many samples. A significant number of accessions are internal distributions to WorldVeg regional offices, and in collaboration with partner institutions, the material has been used in breeding programs. New open-pollinated varieties have been released in Uzbekistan, Tanzania, and Mali through selection based on local trait preferences (Table 4 ).

Table 4 . List of eggplant and African eggplant varieties released in Uzbekistan, Tanzania, and Mali based on WorldVeg germplasm.
Utilization of Eggplant Germplasm in Breeding
Screening of available accessions for targeted traits (evaluation) and morphological description of the accessions (characterization) are key issues for the breeding process. At the WorldVeg a large number of commercial cultivars, landraces, and germplasm have thus been examined to identify desired genotypes for use in eggplant breeding programs or for recommending to private sector seed companies and other partner institutions. Standardized descriptors included characters both for germination, the vegetative phase, inflorescence descriptors, and fruit and seed traits, respectively (Table 5 ).

Table 5 . A complete list of standard descriptors for eggplants used at the World Vegetable Center ( AVGRIS, 2017 ).
Large variation in yield parameters and in fruit quality parameters have been documented in the collection (Figures 2 , 3 ). Such data have been compiled over years and can be retrieved from AVGRIS, the World Vegetable Center genebank database system (2017). Among the 1,308 accessions of S. melongena that have been characterized, green and purple fruits were predominant, and could be found in 47 and 38% of the total number of accessions, respectively. Slightly longer than broad, and as long as broad, were the prevalent shapes of the accessions, with 31.1 and 18.7%, respectively. Similarly, huge diversity was found among 98 accessions belong to S. melongena, S. aethiopicum , and S. macrocarpon for 16 morpho-agronomic and fruit traits including plant height, flowering time, flower/inflorescence, fruit length and fruit acidity, but weak association was found between among morpho-agronomic and fruit quality descriptors ( Polignano et al., 2010 ). In terms of fruit taste, 26.8% of accessions had a sweet taste, 53.2% had an intermediate taste and some accessions had bitter taste (6.1%). Large variations in fruit dry matter content, total sugar content, and fiber content of the fruit have been determined in a study of 90 selected eggplant genotypes ( AVRDC, 1996 ). The distribution of dry matter, total sugar, and fiber contents ranged from 5.5 to 10.1, 7.0 to 40.1, and 4.7 to 18.1%, respectively. In another study conducted at the WorldVeg, 33 S. melongena accessions and two S. aethiopicum accessions were evaluated for superoxide scavenging and content of total phenolics and ascorbic acid ( Hanson et al., 2006 ). Solanum melongena accessions S00062, S00022, and S. aethiopicum accession S00197 exhibited high antioxidant activity ( Hanson et al., 2006 ).

Figure 2 . Horticultural characteristics of more than 1,300 accessions of Solanum melongena summarized and based on information available in AVGRIS (2017) : (A) Fruit color, (B) Fruit length, (C) Fruit yield per plant, and (D) Fruit taste.

Figure 3 . Different fruit shapes, colors, and sizes of Solanum melongena accessions in the World Vegetable Center germplasm collection.
Accessions with important traits such as early maturity, high yielding, and resistance to biotic stresses have been identified in the WorldVeg germplasm collection (Table 6 ). Based on data from Chen (1998) and the examination of 40 accessions from the WorldVeg collection, among long fruit genotypes, VI045551, VI047333, VI046110, and VI037736 were identified as stable and high yielding (>40 tons per hectare) over spring, summer, and autumn seasons. Accession VI046110 had the highest average yield and the earliest maturing genotype across the three seasons ( AVRDC, 1999 ). In round fruit type, VI046097, VI047332, VI44067, EG233, and EG235 produced the high yields in all three seasons.

Table 6 . Identified eggplant germplasm from the World Vegetable Center collection with useful traits for breeding.
Based on data from AVGRIS (2017) compiled over the years and including 1,300 accessions, only 90 accessions (6.8%) had more than 5,000 g of fruit yield per plant (Figure 2 ). Marketable yields were highly associated with fruit weight and number of fruits per plant. Large diversity in the WorldVeg germplasm collections enabled us to develop several improved eggplant and African eggplant cultivars (Table 3 ). A total of three eggplant varieties have been commercialized in Uzbekistan and three African eggplant varieties have been released in Tanzania and Mali.
More than 200 accessions have been evaluated for resistance to bacterial wilt ( Ralstonia solanacearum ) at the WorldVeg under both greenhouse and field conditions ( AVRDC, 1999 ). Among these, 38 accessions were identified with high levels of resistance. These accessions were retested using root wounding and soil drenching inoculation methods in the greenhouse. Data were summarized from the screening and retest studies, and the most resistant accessions were TS3, VI034885, and TS47 from Malaysia; and TS69, TS87, and TS90 from Indonesia with disease indices <10% under both greenhouse and field conditions.
Resistance to eggplant fruit and shoot borer ( Leucinodes orbonalis Guenee), leafhopper ( Amrasca devastans Distant), and aphids ( Aphis gossypii Glover) have been identified at WorldVeg in separate trials ( AVRDC, 1999 ). Leafhoppers and aphids have piercing mouthparts and suck the sap, especially from the leaves, which leads to yellow spots on the leaves, followed by crinkling, curling, bronzing, and drying (or “hopper burn” from leafhopper), but severe aphid infestations cause young leaves to curl and become deformed ( AVRDC, 1999 ; Ramasamy, 2009 ). Like whiteflies, aphids also produce honeydew, which leads to the development of sooty mold ( Ramasamy, 2009 ). Accessions VI034971, VI035822, and VI035835 were found promising accessions against leafhopper and aphids. Eggplant fruit and shoot borer is an extremely destructive pest, especially in South and Southeast Asia ( Ramasamy, 2009 ). It lays eggs on the foliage and neonate larvae feeds on the tender shoots, boring into the shoots and fruits, resulting in wilting of young shoots, followed by drying; the fruit becomes unfit for marketing and consumption. Total resistance was not found and moderate resistance was found only in one accession, VI047451 ( AVRDC, 1999 ). This was based on typical damage symptoms, wilting of shoots and feeding holes in a wilted shoot, as well as damaged fruit. Overall, these results show that very promising materials for breeding pest tolerant or resistant varieties can be found in the WorldVeg eggplant collection. However, additional race specific screening is needed to find resistant sources for pests where no resistance or limited resistance has been found.
The Way Forward
The food security of many countries relies on crops bred from genetic resources outside their region ( Khoury et al., 2016 ). Therefore, plant genetic resources are a global concern where access and benefit sharing is of paramount importance. Eggplant is an important vegetable crop with a global cultivation area. From the current study we have confirmed that there are critical gaps in global eggplant collections, especially related to crop wild relatives ( Syfert et al., 2016 ). We have listed more than 35 wild species conserved in germplasm collections, but for many other eggplant wild relatives no accessions are conserved in genebanks; in addition, there still might be undiscovered crop wild relatives. Genetic diversity in wild relatives is much higher than in cultivated eggplant ( Vorontsova et al., 2013 ) and could be valuable sources for resistance to biotic and abiotic stresses ( Daunay and Hazra, 2012 ). To date, a limited number of wild relatives have used in eggplant breeding ( Rotino et al., 2014 ) and commercial varieties containing wild relative introgressions are not yet available. To move forward, screening for abiotic and biotic stresses in wild relatives should be intensified and broadened for identification of valuable germplasm accessions for breeding improved eggplant varieties. This information, combined with genomics studies for the detection of genes and QTLs of agronomic importance and their associated markers, will be of great utility in eggplant breeding, as has been demonstrated in some association mapping studies ( Cericola et al., 2014 ; Portis et al., 2015 ). Recent reviews of the development in eggplant is provided by Frary and Doganlar (2013) and Gramazio et al. (in press) .
From a utilization point of view, core collections could be established and stakeholders should work together for the development of the next generation of eggplant varieties that can meet the challenges of the present and the future.
Author Contributions
DT compiled the major parts of the text; SS contributed with text on genetic resources; JP contributed with text on eggplant wild relatives; YC contributed with reviewing databases; MR and TW contributed with inputs on eggplant taxonomy and breeding.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Funding for the World Vegetable Center's general research activities is provided by core donors: Republic of China (Taiwan), UK aid, United States Agency for International Development (USAID), Australian Centre for International Agricultural Research (ACIAR), Germany, Thailand, Philippines, Korea, and Japan. In addition we like to thank Global Crop Diversity Trust for contribution to meetings and to this open-access publication.
AVGRIS (2017). The AVRDC Vegetable Genetic Resources Information System . Available online at: https://avrdc.org/our-work/managing-germplasm
AVRDC (1996). AVRDC 1995 Report . Asian Vegetable Research and Development Center, Tainan, 42–45.
AVRDC (1999). AVRDC 1998 Report . Asian Vegetable Research and Development Center, Tainan, 32–36.
Braga, P. C., Lo Scalzo, R., dal Sasso, M., Lattuada, N., Greco, V., and Fibiani, M. (2016). Characterization and antioxidant activity of semi-purified extracts and pure delphinine-glycosides from eggplant peel ( Solanum melongena L.) and allied species. J. Funct. Foods 20, 411–421. doi: 10.1016/j.jff.2015.10.032
CrossRef Full Text | Google Scholar
Bukenya, Z. R., and Carasco, J. F. (1994). Biosystematic study of Solanum macrocarpon - S. dasyphyllum complex in Uganda and relations with S. linnaeanum . J. East Afr. Agric. For. 59, 187–204.
Google Scholar
Cao, G., Sofic, E., and Prior, R. L. (1996). Antioxidant capacity of tea and common vegetables. J. Agric. Food Chem. 44, 3426–3431. doi: 10.1021/jf9602535
Castañeda-Álvarez, N. P., Khoury, C. K., Achicanoy, H. A., Bernau, V., Dempewolf, H., Eastwood, R. J., et al. (2016). Global conservation priorities for crop wild relatives. Nat. Plants 2:16022. doi: 10.1038/nplants.2016.22
PubMed Abstract | CrossRef Full Text | Google Scholar
Cericola, F., Portis, E., Lanteri, S., Toppino, L., Barchi, L., Acciarri, N., et al. (2014). Linkage disequilibrium and genome-wide association analysis for anthocyanin pigmentation and fruit color in eggplant. BMC Genomics 15:896. doi: 10.1186/1471-2164-15-896
Cericola, F., Portis, E., Toppino, L., Barchi, L., Acciarri, N., Ciriaci, T., et al. (2013). The population structure and diversity of eggplant from Asia and the Mediterranean basin. PLoS ONE 8:e73702. doi: 10.1371/journal.pone.0073702
Chen, N. C. (1998). Evaluation of Elite Eggplant Cultivars. AVRDC 1998 Report , 30–32. World Vegetable Center, Tainan.
Chiarini, F. E., Moreno, N. C., Barboza, G. E., and Bernardello, G. (2010). Karyotype characterization of Andean Solanoideae (Solanaceae). Caryologia 63, 278–291. doi: 10.1080/00087114.2010.589738
Daunay, M. C., and Hazra, P. (2012). “Eggplant,” in Handbook of Vegetables , eds K. V. Peter and P. Hazra (Houston, TX: Studium Press), 257–322.
Daunay, M. C., Lester, R. N., and Ano, G. (2001). “Eggplant,” in Tropical Plant Breeding , eds A. Charrier, A. Jacquot, M. Hamon, and D. Nicolas (Montpellier: Science Publishers), 199–222.
Docimo, T., Francese, G., Ruggiero, A., Batelli, G., De Palma, M., Bassolino, L., et al. (2016). Phenylpropanoids accumulation in eggplant fruit: characterization of biosynthetic genes and regulation by a MYB transcription factor. Front. Plant Sci. 6:1233. doi: 10.3389/fpls.2015.01233
FAO (2010). The Second Report on the State of the World's Plant Genetic Resources for Food and Agriculture . Food and Agricultural Organization, Rome.
FAO (2014). FAOSTAT Production Databases . Available online at: http://www.faostat.fao.org (Accessed January 30, 2017).
Frary, A., and Doganlar, S. (2013). “Eggplants,” in Genetics, Genomics and Breeding of Peppers and Eggplants , eds B. C. Kang and C. Kole (Boca Raton, FL: CRC Press; Taylor & Francis Group), 116–143. doi: 10.1201/b14541-8
Frary, A., Doganlar, S., and Daunay, M. C. (2007). “Eggplant,” in Vegetables SE - 9, Genome Mapping and Molecular Breeding in Plants , ed C. Kole (Berlin: Springer), 287–313. doi: 10.1007/978-3-540-34536-7_9
Frodin, D. G. (2004). History and concepts of big plant genera. Taxon 53, 753–776. doi: 10.2307/4135449
GBIF (2017). The Global Biodiversity Information Facility . Available online at: http://www.gbif.org (Accessed January 20, 2017).
GENESYS (2017). The Global Gateway to Genetic Resources . Available online at: https://www.genesys-pgr.org (Accessed January 20, 2017).
Gramazio, P., Prohens, J., Plazas, M., Mangini, G., Herraizi, F., Garcia-Fortea, E., et al. (in press). Genomic tools for the enhancement of vegetable crops: a case in eggplant. Notulae Botan. Horti Agrobot. Cluj-Napoca .
Hanson, P. M., Yang, R., Tsou, S. C., Ledesma, D., Engle, L., and Leeb, T. (2006). Diversity in eggplant ( Solanum melongena ) for superoxide scavenging activity, total phenolics, and ascorbic acid. J. Food Comp. Anal. 19, 594–600. doi: 10.1016/j.jfca.2006.03.001
Harlan, J. R., and de Wet, J. M. J. (1971). Toward a rational classification of cultivated plants. Taxon 20, 509–517. doi: 10.2307/1218252
Hurtado, M., Vilanova, S., Plazas, M., Gramazio, P., Fonseka, H. H., Fonseka, R., et al. (2012). Diversity and relationships of eggplants from three geographically distant secondary centers of diversity. PLoS ONE 7:e41748. doi: 10.1371/journal.pone.0041748
Khoury, K. C., Achinacoy, H. A., Bjorkman, A. D., Navarro-Racines, C., Guarino, L., Flores-Palacios, X., et al. (2016). Origins of food crops connects countries worldwide. Proc. R. Soc. B 283:20160792. doi: 10.1098/rspb.2016.0792
Knapp, S., Vorontsova, M. S., and Prohens, J. (2013). Wild relatives of the eggplant ( Solanum melongena L.: Solanaceae): new understanding of species names in a complex group. PLoS ONE 8:e57039. doi: 10.1371/journal.pone.0057039
Kouassi, B., Prohens, J., Gramazio, P., Kouassi, A. B., Vilanova, S., Galán-Ávila, A., et al. (2016). Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant ( Solanum melongena ). Sci. Hort. 213, 199–207. doi: 10.1016/j.scienta.2016.10.039
Lester, R. N. (1986). Taxonomy of scarlet eggplants, Solanum aethiopicum L. Acta Hort. 182, 125–132. doi: 10.17660/ActaHortic.1986.182.15
Lester, R. N., and Daunay, M. C. (2003). Diversity of African vegetable Solanum species and its implications for a better understanding of plant domestication. Schriften Genetischen Ressour. 22, 137–152.
Lester, R. N., and Hasan, S. M. (1991). “Origin and domestication of the brinjal egg-plant, Solanum melongena , from S. incanum , in Africa and Asia,” in Solanaceae III: Taxonomy, Chemistry, Evolution , eds J. G. Hawkes, R. N. Lester, M. Nee, and N. Estrada (Kew: Royal Botanic Gardens), 369–387.
Lester, R. N., Jaeger, P. M., and Child, A. (2011). Solanum in Africa . Birmingham: Celia Lester.
Lester, R. N., and Niakan, L. (1986). “Origin and domestication of the scarlet eggplant, Solanum aethiopicum , from S. anguivi in Africa,” in Solanaceae: Biology and Systematics , ed W. G. D'Arcy (New York, NY: Columbia University Press), 433–456.
Lester, R. N., and Thitai, G. N. W. (1989). Inheritance in Solanum aethiopicum , the scarlet eggplant. Euphytica 40, 67–74.
Levin, R. A., Myers, N. R., and Bohs, L. (2006). Phylogenetic relationships among the “spiny solanums” ( Solanum subgenus Leptostemonum , Solanaceae). Am. J. Bot. 93, 157–169. doi: 10.3732/ajb.93.1.157
Maundu, P., Achigan-Dako, E., and Morimoto, Y. (2009). “Biodiversity of African vegetables,” in African Indigenous Vegetables in Urban Agriculture , eds C. M. Shackleton, M. W. Pasquini, and A. W. Drescher (London: Earthscan), 65–104.
Medakker, A., and Vijayaraghavan, V. (2007). “Successful commercialization of insect-resistant eggplant by a public–private partnership: reaching and benefiting resource-poor farmers” in Intellectual Property Management in Health and Agricultural Innovation: A Handbook of Best Practices , eds A. Krattiger, R. T. Mahoney, L. Nelsen, J. A. Thomson, A. B. Bennett, K. Satyanarayana, G. D. Graff, C. Fernandez, and S. P. Kowalski (Oxford: MIHR and Davis; PIPRA). Available online at: www.ipHandbook.org
Mennella, G., Lo Scalzo, R., Fibiani, M., D'Alessandro, A., Francese, G., Toppino, L., et al. (2012). Chemical and bioactive quality traits during fruit ripening in eggplant ( S. melongena L.) and allied species. J. Agric. Food Chem. 60, 11821–11831. doi: 10.1021/jf3037424
Meyer, R. S., Karol, K. G., Little, D. P., Nee, M. H., and Litt, A. (2012). Phylogeographic relationships among Asian eggplants and new perspectives on eggplant domestication. Mol. Phylogenet. Evol. 63, 685–701. doi: 10.1016/j.ympev.2012.02.006
Meyer, R. S., Whitaker, B. D., Little, D. P., Wu, S. B., Kennelly, E. J., Long, C. L., et al. (2015). Parallel reductions in phenolic constituents resulting from the domestication of eggplant. Phytochemistry 115, 194–206. doi: 10.1016/j.phytochem.2015.02.006
Nee, M. (1999). “Synopsis of Solanum in the New World,” in Solanaceae IV: Advances in Biology and Utilization , eds M. Nee, D. E. Symon, R. N. Lester, and J. P. Jessop (Kew, VIC: Royal Botanic Gardens), 285–333.
Plazas, M., Andújar, I., Vilanova, S., Gramazio, P., Herraiz, F. J., et al. (2014a). Conventional and phenomics characterization provides insight into the diversity and relationships of hypervariable scarlet ( Solanum aethiopicum L.) and gboma ( S. macrocarpon L.) eggplant complexes. Front. Plant Sci. 5:318. doi: 10.3389/fpls.2014.00318
Plazas, M., Andújar, I., Vilanova, S., Hurtado, M., Gramazio, P., Herraiz, F. J., et al. (2013). Breeding for chlorogenic acid content in eggplant: interest and prospects. Not. Bot. Horti. Agrobot. 41, 26–35. doi: 10.15835/nbha4119036
Plazas, M., Prohens, J., Cuñat, A. N., Vilanova, S., Gramazio, P., Herraiz, F. J., et al. (2014b). Reducing capacity, chlorogenic acid content and biological activity in a collection of scarlet ( Solanum aethiopicum ) and gboma ( S. macrocarpon ) eggplants. Int. J. Mol. Sci. 15, 17221–17241. doi: 10.3390/ijms151017221
Plazas, M., Vilanova, S., Gramazio, P., Rodríguez-Burruezo, A., Fita, A., Herraiz, F. J., et al. (2016). Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hort. Sci. 141, 34–44.
Polignano, G., Uggenti, P., Bisignano, V., and Della Gatta, C. (2010). Genetic divergence analysis in eggplant ( Solanum melongena L.) and allied species. Genet. Resour. Crop Evol. 57, 171–181. doi: 10.1007/s10722-009-9459-6
Portis, E., Cericola, F., Barchi, L., Toppino, L., Acciarri, N., Pulcini, L., et al. (2015). Association mapping for fruit, plant and leaf morphology traits in eggplant. PLoS ONE 10:e0135200. doi: 10.1371/journal.pone.0135200
Prohens, J., Blanca, J. M., and Nuez, F. (2005). Morphological and molecular variation in a collection of eggplant from a secondary center of diversity: implications for conservation and breeding. J. Am. Soc. Hort. Sci. 130, 54–63.
Prohens, J., Plazas, M., Raigón, M. D., Seguí-Simarro, J. M., Stommel, J. R., and Vilanova, S. (2012). Characterization of interspecific hybrids and first backcross generations from crosses between two cultivated eggplants ( Solanum melongena and S. aethiopicum Kumba group) and implications for eggplant breeding. Euphytica 186, 517–538. doi: 10.1007/s10681-012-0652-x
Raigón, M. D., Prohens, J., Muñoz-Falcón, J. E., and Nuez, F. (2008). Comparison of eggplant landraces and commercial varieties for fruit content of phenolics, minerals, dry matter and protein. J. Food Comp. Anal. 21, 370–376. doi: 10.1016/j.jfca.2008.03.006
Ramasamy, S. (2009). Insect and Mite Pests on Eggplant: A Field Guide for Indentification and Management . AVRDC – The World Vegetable Center, Shanhua, Taiwan. AVRDC Publication No. 09-729.
Ranil, R. H., Prohens, J., Aubriot, X., Niran, H. M., Plazas, M., Fonseka, R. M., et al. (2017). Solanum insanum L. (subgenus Leptostemonum Bitter, Solanaceae), the neglected wild progenitor of eggplant ( S. melongena L.): a review of taxonomy, characteristics and uses aimed at its enhancement for improved eggplant breeding. Genet. Resour. Crop. Evol . doi: 10.1007/s10722-016-0467-z. [Epub ahead of print].
Rotino, G. L., Perri, E., Acciarri, N., Sunseri, F., and Arpaia, S. (1997). Development of eggplant varietal resistance to insects and diseases via plant breeding. Adv. Hort. Sci. 11, 193–201.
Rotino, G. L., Sala, T., and Toppino, L. (2014). “Eggplant,” in Alien Gene Transfer in Crop Plants, Vol. 2 , eds A. Pratap and J. Kumar (New York, NY: Springer), 381–409. doi: 10.1007/978-1-4614-9572-7_16
Schippers, R. R. (2000). African Indigenous Vegetables: An Overview of the Cultivated Species . Natural Resources Institute/ACP-EU Technical Centre for Agricultural and Rural Cooperation, Chatham, UK.
SGSV (2017). The Seed Portal of the Svalbard Global Seed Vault . Available online at http://www.nordgen.org/sgsv (Accessed January 20, 2017).
Stern, S., Agra, M. F., and Bohs, L. (2011). Molecular delimitation of clades within New World species of the “spiny solanums” ( Solanum subg. Leptostemonum ). Taxon 60, 1429–1441.
Stommel, J. R., Whitaker, B. D., Haynes, K. G., and Prohens, J. (2015). Genotype × environment interactions in eggplant for fruit phenolic acid content. Euphytica 205, 823–836. doi: 10.1007/s10681-015-1415-2
Syfert, M. M., Castañeda-Álvarez, N. P., Khoury, C. K., Särkinen, T., Sosa, C. C., Achicanoy, H. A., et al. (2016). Crop wild relatives of the brinjal eggplant ( Solanum melongena ): poorly represented in genebanks and many species at risk of extinction. Am. J. Bot. 103, 635–651. doi: 10.3732/ajb.1500539
Vavilov, N. I. (1951). The Origin, Variation, Immunity and Breeding of Cultivated Plants. Transl. by K. Start. Waltham, MA: Chronica Botanica.
Vilanova, S., Manzur, J. P., and Prohens, J. (2012). Development and characterization of genomic simple sequence repeat markers in eggplant and their application to the study of diversity and relationships in a collection of different cultivar types and origins. Mol. Breed. 30, 647–660. doi: 10.1007/s11032-011-9650-2
Vorontsova, M. S., and Knapp, S. (2012). A new species of Solanum (Solanaceae) from South Africa related to the cultivated eggplant. PhytoKeys 8, 1–11. doi: 10.3897/phytokeys.8.2462
Vorontsova, M. S., and Knapp, S. (2016). A revision of the spiny solanums, Solanum subgenus Leptostemonum (Solanaceae) in Africa and Madagascar. Syst. Bot Monogr. 99, 1–436.
Vorontsova, M. S., Stern, S., Bohs, L., and Knapp, S. (2013). African spiny Solanum subgenus Leptostemonum , Solanaceae): a thorny phylogenetic tangle. Bot. J. Linn. Soc. 173, 176–193. doi: 10.1111/boj.12053
Weese, T., and Bohs, L. (2010). Eggplant origins: out of Africa, into the Orient. Taxon 59, 49–56. doi: 10.2307/27757050
Keywords: conservation, crop wild relatives, diversity, plant genetic resources, Solanum melongena , Solanum aethiopicum , Solanum macrocarpon
Citation: Taher D, Solberg SØ, Prohens J, Chou Y, Rakha M and Wu T (2017) World Vegetable Center Eggplant Collection: Origin, Composition, Seed Dissemination and Utilization in Breeding. Front. Plant Sci . 8:1484. doi: 10.3389/fpls.2017.01484
Received: 10 May 2017; Accepted: 10 August 2017; Published: 25 August 2017.
Reviewed by:
Copyright © 2017 Taher, Solberg, Prohens, Chou, Rakha and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY) . The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Svein Ø. Solberg, [email protected]
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
Field Performance of Bt Eggplants ( Solanum melongena L.) in the Philippines: Cry1Ac Expression and Control of the Eggplant Fruit and Shoot Borer ( Leucinodes orbonalis Guenée)
Desiree m. hautea.
1 Institute of Plant Breeding/CSC, College of Agriculture, University of the Philippines Los Baños, College, Laguna, 4031, Philippines
Lourdes D. Taylo
Anna pauleen l. masanga, maria luz j. sison, josefina o. narciso, reynaldo b. quilloy, randy a. hautea.
2 International Service for the Acquisition of Agri-Biotech Applications, Los Baños, Laguna, 4030, Philippines
Frank A. Shotkoski
3 International Programs, Cornell University, Ithaca, New York, 14853, United States of America
Anthony M. Shelton
4 Department of Entomology, Cornell/NYSAES, Geneva, New York, 14456, United States of America
Conceived and designed the experiments: DMH RAH JON LDT MLJS FAS AMS. Performed the experiments: JON RBQ LDT MLJS APLM DMH RAH. Analyzed the data: LDT APLM DMH RAH. Wrote the paper: DMH LDT APLM RAH FAS AMS. Developed the breeding materials: JON LDT RBQ DMH RAH. Supervised the field trials: DMH RAH RBQ LDT FAS. Prepared application and obtained all relevant permits for confined field trials: DMH JON LDT RAH.
Associated Data
Data are available from Dryad (doi: 10.5061/dryad.ks131 ).
Plants expressing Cry proteins from the bacterium, Bacillus thuringiensis (Bt), have become a major tactic for controlling insect pests in maize and cotton globally. However, there are few Bt vegetable crops. Eggplant ( Solanum melongena ) is a popular vegetable grown throughout Asia that is heavily treated with insecticides to control the eggplant fruit and shoot borer, Leucinodes orbonalis (EFSB). Herein we provide the first publicly available data on field performance in Asia of eggplant engineered to produce the Cry1Ac protein. Replicated field trials with five Bt eggplant open-pollinated (OP) lines from transformation event EE-1 and their non-Bt comparators were conducted over three cropping seasons in the Philippines from 2010–2012. Field trials documented levels of Cry1Ac protein expressed in plants and evaluated their efficacy against the primary target pest, EFSB. Cry1Ac concentrations ranged from 0.75–24.7 ppm dry weight with the highest in the terminal leaves (or shoots) and the lowest in the roots. Cry1Ac levels significantly increased from the vegetative to the reproductive stage. Bt eggplant lines demonstrated excellent control of EFSB. Pairwise analysis of means detected highly significant differences between Bt eggplant lines and their non-Bt comparators for all field efficacy parameters tested. Bt eggplant lines demonstrated high levels of control of EFSB shoot damage (98.6–100%) and fruit damage (98.1–99.7%) and reduced EFSB larval infestation (95.8–99.3%) under the most severe pest pressure during trial 2. Moths that emerged from larvae collected from Bt plants in the field and reared in their Bt eggplant hosts did not produce viable eggs or offspring. These results demonstrate that Bt eggplant lines containing Cry1Ac event EE-1 provide outstanding control of EFSB and can dramatically reduce the need for conventional insecticides.
Introduction
Since their introduction in 1996, maize and cotton expressing insecticidal proteins derived from the soil bacterium Bacillus thuringiensis (Bt) have been widely adopted and in 2014 were planted on 78.8 million ha in 28 countries predominantly by resource-poor farmers [ 1 ]. Bt crops are another form of host plant resistance, the foundation for integrated pest management (IPM) programs [ 2 ]. Several major maize and cotton pests have been successfully controlled, and insecticide use on them has been substantially reduced throughout most adopting countries [ 3 ]. Unfortunately, the development of Bt crops has been limited to major commodity crops (maize, cotton, and soybean) and not fruit and vegetables, except sweet corn. This situation is especially unfortunate since fruit and vegetables, when taken together, receive more insecticides than maize, cotton and rice combined [ 4 ].
Eggplant, Solanum melongena L. (also known as brinjal and aubergine) is one of the most important, inexpensive and popular vegetable crops grown and consumed in Asia. In the Philippines, eggplant production accounts for more than 30.0% of the total volume of production of the most important vegetables in the country [ 5 ]. Eggplant production provides an important source of cash income, particularly for small, resource-poor farmers. The biggest constraint to eggplant production throughout Asia is the chronic and widespread infestation by the eggplant fruit-and-shoot borer (EFSB), Leucinodes orbonalis Guenée [ 6 ]. The larvae damage eggplant by boring into the petiole and midrib of leaves and tender shoots resulting in wilting and desiccation of stems. Flowers are also fed upon resulting in flower drop or misshapen fruits. The most serious economic damage caused by EFSB is to the fruit by producing holes, feeding tunnels and frass (or larval excrement) that make the fruit unmarketable and unfit for human consumption. At high pest pressure, EFSB damage in the Philippines results in yield loss of up to 80.0% of the crop [ 7 ]. Surveys of eggplant farmers in the major eggplant growing provinces of the Philippines [ 7 – 11 ] revealed that almost all of them use chemical insecticides to control EFSB because other control measures such as manual removal of EFSB-damaged fruits and wilted shoots, use of biological control arthropods and pheromone traps [ 12 ] have proven ineffective, impractical and expensive. Eggplant farmers in the Philippines employ frequent applications (20–72 times for 5–6 months/season) of mixtures of insecticides to control EFSB, which increase production costs and pose risks to human health and the environment. Studies conducted in Sta. Maria, Pangasinan [ 8 , 13 , 14 ] showed frequent use of broad-spectrum insecticides including profenofos, triazophos, chlorpyrifos, cypermethrin, and malathion. Residues of these insecticides were detected in the soil of eggplant farms and in harvested fruits [ 14 ]. Farmers and farm workers in the study attributed various ailments such as skin irritation, redness of the eyes, muscle pains and headaches to exposure to these pesticides.
After more than 40 years, conventional breeding has not produced any commercial variety of eggplant conferring high level of resistance to the EFSB [ 15 ]. Therefore, efforts became focused on developing Bt eggplant that expresses the same Cry1Ac protein as the cotton event MON531, which has been approved by regulatory agencies in many countries [ 16 – 18 ]. MON531 has been bred into cotton varieties that have been on the global market for almost 20 years with no verifiable report of any adverse effect on human health or the environment. The modified gene used in MON531 encodes an amino acid sequence that is 99.4% identical to the naturally occurring microbial Cry1Ac protein [ 19 , 20 ].
Maharashtra Hybrid Seeds Co. Pvt. Ltd. (Mahyco) inserted the cry1Ac gene under the control of the constitutive 35S CaMV promoter into eggplant to control feeding damage caused by EFSB [ 21 ]. The transformation event designated as 'EE-1' was introgressed into eggplant varieties and hybrids in India, Bangladesh and the Philippines [ 22 , 23 ]. In 2009, although the Indian biosafety regulatory agency gave biosafety approval to Mahyco event EE-1, the Ministry of the Environment and Forests placed a moratorium on its cultivation in India [ 24 ] that remains in effect as of May 2016. In 2013, four Bt eggplant varieties containing the same EE-1 event were conditionally approved for cultivation in Bangladesh. These were grown on 20 fields in 2014 and the number increased to 108 farms in 2015 ( https://bteggplant.wordpress.com/2015/08/11/speech-by-dr-md-rafiqul-islam-mondal-director-general-bari/ ). In the Philippines, event EE-1 was introgressed into selected EFSB-susceptible eggplant open-pollinated (OP) varieties through conventional backcrossing coupled with diagnostic EE-1 event-specific PCR and a cry1Ac gene strip assay [ 25 ]. Five promising advanced Bt OP lines, developed by the University of the Philippines Los Baños, were selected for Confined Field Trial testing in selected eggplant growing areas of the country.
The studies presented in this report contain the first data on Bt eggplant for control of EFSB in Asia to be submitted to a peer-reviewed journal. The studies were conducted with the following objectives: (1) to determine the expression levels of Cry1Ac protein in Bt eggplant OP lines; and (2) to evaluate the field efficacy of the EE-1 event in Bt eggplant OP lines against field populations of EFSB. Results of these studies will be used to generate crucial information for selecting the best EFSB-resistant Bt eggplant OP lines for market release in the Philippines.
Bt Cry1Ac protein expression in different plant parts in Bt Eggplant OP lines
Significant differences were detected in Cry1Ac protein expression among the different plant parts in all Bt eggplant OP lines grown for two seasons ( Fig 1 , S1 Table ). The highest levels of Cry1Ac protein were detected in the terminal leaves, with decreasing levels of expression in the flowers, fruits, stem and roots. Results of the gene strip test of the non-Bt eggplant comparators (near-isoline counterparts and check) were negative and the quantitative ELISA values were below the limit of quantitation (LOQ = 0.125) of the assay used.

Mean ± SEM Cry1Ac protein concentration in terminal leaves, flowers, fruits, stem, and roots of five (5) Bt eggplant OP lines. n = 4 per plant part/Bt line with 5–10 sample plants per replicate plot. Confined field trial1 (wet/off season) and trial 2 (dry season), CY 2010–2011, Pangasinan, Philippines.
Terminal Leaves
The Cry1Ac protein expressed in terminal leaves ranged from 18.32–24.87 ppm dry weight (DW) in trial 1, and 20.40–21.83 ppm DW in trial 2. Line M4 expressed significantly less Cry1Ac protein than line D2 in trial 1; however there were no other significant differences in expression levels among the Bt eggplant lines in either trial.
There were no significant differences detected in Cry1Ac protein content among the Bt eggplant lines in both trials. Cry1Ac protein content in the flowers ranged from 10.17–16.33 ppm DW in trial 1, and from 14.34–17.57 ppm DW in trial 2.
Fruits (flesh and skin)
There were no significant differences in Cry1Ac protein expression in either the fruit flesh or the skin among the Bt eggplant lines tested in both trials. However, the widest range of variation was observed between the two trials. In trial 1, the fruit flesh contained higher levels of Cry1Ac protein at 9.00–16.23 ppm DW and the fruit skin ranged from 8.82–13.42 ppm DW, but much lower levels of Cry1Ac protein were detected in both the flesh and skin in trial 2 (3.02–9.47 and 2.61–7.18 ppm DW, respectively).
There were no significant differences in Cry1Ac protein expression in the stem among all Bt eggplant lines tested. The stem contained Cry1Ac protein concentration of 2.75–5.22 ppm DW in trial 1 and 5.00–7.02 ppm DW in trial 2.
The roots contained the lowest levels of Cry1Ac protein. The mean Cry1Ac protein concentration (1.8 ppm DW) was similar in both trials. There were no significant differences in Cry1Ac protein expression in the roots among the Bt eggplant lines tested. The highest level of Cry1Ac protein expressed in the roots was 2.64 ppm DW.
Cry1Ac protein expression in terminal leaves at different growth stages in Bt Eggplant OP lines
Significant differences in Cry1Ac protein expression were detected in terminal leaves across three growth stages of eggplant development in all Bt eggplant OP lines in both trials ( Fig 2 , S2 Table ). The observed pattern of Cry1Ac protein expression generally increased from the vegetative stage to the reproductive stage; then at the late reproductive stage the levels slightly decreased in trial 1 but increased in trial 2. Higher concentrations of Cry1Ac protein were detected in trial 2 at the vegetative stage (18.69–19.22 ppm DW) and late reproductive stage (22.32–23.54 ppm DW) compared with amounts detected at the same growth stages during trial 1. Significant differences were observed among the Bt eggplant lines in the amount of Cry1Ac protein expressed during the vegetative and reproductive stages in trial 1. Bt eggplant line M4 showed the lowest level of Cry1Ac protein expression among the lines tested. However, no significant differences were observed among the Bt eggplant lines at all growth stages in trial 2.

Mean ± SEM Cry1Ac protein concentration in the terminal leaves of 5 Bt eggplant OP lines at the vegetative, reproductive and late reproductive stages. n = 4 per growth stage/line with 5–10 plants per replicate plot. Confined field trial 1 (wet/off season) and trial 2 (dry season), CY 2010–2011, Pangasinan, Philippines.
Control of EFSB by Bt Eggplant OP lines
Under natural field infestations, the efficacy against EFSB of Bt eggplant lines and the non-Bt comparators (near-isoline counterparts and check variety) were evaluated for three seasons (trials 1–3) based on the following parameters: % EFSB-damaged shoots, % EFSB-damaged fruits and number of EFSB larvae in fruits. Throughout the sampling/harvest periods, Bt eggplant lines consistently demonstrated a lower percentage of EFSB-damaged shoots ( Fig 3 , S3 Table ), % EFSB- damaged fruits ( Fig 4 , S4A to S4C Table ) and number of EFSB larvae in fruits ( Fig 5 , S4A to S4C Table ) compared to the conventionally-bred non-Bt eggplant comparators.

Mean ± SEM percentage (%) EFSB-damaged shoots in Bt lines and their non-Bt eggplant comparators at different sampling periods. n = 4 per entry with 16 plants per replicate plot. Confined field trials 1 and 3 (wet/off season) and trial 2 (dry season), CY 2010–2012, Pangasinan, Philippines.

Mean ± SEM percentage (%) EFSB-damaged fruits in Bt lines and their non-Bt eggplant comparators at different harvest periods. n = 4 per entry with 16 plants per replicate plot. Confined field trials 1 and 3 (wet/off season) and trial 2 (dry season), CY 2010–2012, Pangasinan, Philippines.

Mean ± SEM number of EFSB larvae in damaged fruits of Bt lines and their non-Bt eggplant comparators at different harvest periods. n = 4 per entry with 16 plants per replicate plot. Confined field trials 1 and 3 (wet/off season) and trial 2 (dry season), CY 2010–2012, Pangasinan, Philippines.
Significant differences among entries ( P = <0 . 0001 ) were detected in all parameters measured in the three trials. Results of paired mean comparison by contrast for all parameters and corresponding level of control (% efficacy) relative to non-Bt eggplant are presented in Table 1 . Highly significant differences were consistently detected between Bt eggplant lines and their corresponding non-Bt eggplant comparators for all parameters in every trial. Comparisons between Bt eggplant lines and non-Bt eggplant comparators showed significantly lower shoot and fruit damage and fewer surviving EFSB larvae in fruits in the Bt eggplant lines tested. Bt eggplant demonstrated 97.7–100% and 96.0–100% control of EFSB shoot and fruit damage, respectively, except in Bt line M4 in trial 1. This line had slightly lower % efficacy for shoot damage (95.3%) and fruit damage (94.1%) but these levels were significanly much better than any of the the non-Bt comparators. Control of EFSB larval infestation in Bt eggplant lines ranged from 88.4–100%, with most lines showing > 96.0% control, except for Bt M1 and M8 in trial 1. Nevertheless, the levels of control of EFSB larval infestation in M1 and M8 were still far better compared to any of the non-Bt comparators tested.
1 Mean comparison by contrast (PROC MIXED in SAS);
** highly significant at 1% probability level; mean of 4 replicates;
2 Bt = eggplant containing event ‘EE-1’; NBt = non-Bt eggplant near-isolines and commercial check;
3 Mean of 10 weekly observation periods;
4 Mean of total harvest periods: Trial 1 (9 harvests); Trial 2 (13 harvests); Trial 3 (9 harvests)
5 Mean of total harvest periods: Trial 1 (9 harvests); Trial 2 (13 harvests); Trial 3 (9 harvests)
6 %Efficacy = (1-Bt/non-Bt) x 100
Seasonal variation in field damage was also observed between Bt eggplant lines and their non-Bt comparators ( Table 1 ). Trials 1 and 3 were conducted during the wet/off planting season when most of the surrounding annual crop was rice. Trial 2 was conducted during the dry season, when eggplants are more widely grown in Pangasinan. Of the three trials conducted, the highest pest pressure was recorded during trial 2 as evidenced by the highest percentages of plant damage and number of insects observed. During trial 2, the highest mean % EFSB-damaged shoots (41.58%), % EFSB-damaged fruits (93.08%), and number of surviving EFSB larvae (16.15 larvae/plot/harvest) were recorded in the non-Bt eggplant comparators. Under such severe pest pressure, the Bt eggplant lines showed <1% EFSB shoot damage, <2% fruit damage and fewer EFSB larvae (<1 larva/plot/harvest).
Survival and Fecundity of EFSB in Bt Eggplant OP lines
EFSB larvae were collected from plants of Bt eggplant lines and non-Bt comparators and brought to the Entomology laboratory and reared continuously in their respective hosts. The results showed that very few EFSB larvae were collected in all Bt eggplant compared with the non-Bt eggplant plants sampled ( Table 2 ). Of the total (27) EFSB larvae reared from Bt eggplant plants, less than half (11/27) emerged as adults and almost half (5/11) of the moths were weak and died before mating. Only six adults were able to mate successfully. However, no viable eggs and offspring resulted from any paired matings involving either male or female EFSB adults collected and reared in the Bt eggplants ( Table 3 ). In contrast, a high percentage (97.3%) of EFSB larvae collected from the non-Bt plants successfully emerged as adults, mated and produced many viable eggs and young larvae.
1 EFSB larvae collected from Bt and non-Bt hosts from larvae collected from trial 2 in Pangasinan and reared continuously on respective hosts in the IPB Entomology P2 Laboratory
2 Bt = eggplant containing event EE-1; NBt = non-Bt eggplant counterpart genotypes
3 Total larval counts collected in 5 Bt OP lines and 2 non-Bt counterparts; 4 reps and 7 harvests
4 Mean of 5 Bt OP lines and 2 non-Bt OP counterpart lines; 4 reps and 7 harvests
5 Relative % = (Total Bt)/(Total (Bt + Non-Bt)) x 100; (Total Non-Bt)/(Total (Bt + Non-Bt)) x 100
1 Bt = eggplant containing event EE-1; NBt = non-Bt eggplant counterpart genotypes
2 Paired matings included all surviving adults collected from Bt hosts reared continuously on same hosts; paired mating of the control were representative samples of surviving adults collected from non-Bt hosts reared continuously on the same host
Spatio-Temporal Expression of Cry1Ac Protein in Bt Eggplant Lines
Recent reviews [ 18 , 26 , 27 ] of Bt crops engineered to express δ-endotoxin proteins cited numerous reports indicating that the expression of Cry proteins vary with plant parts, plant age, genotypes and environmental conditions. To provide the greatest benefits, Cry proteins should be expressed in sufficient quantities to provide high levels of protection to appropriate plant parts and at the stage of growth when the target insect pest pressure is most severe. In this study, significant differences were detected in the amount of Cry1Ac expressed in different plant parts: terminal leaves > flowers > fruits > stem > roots. Cry1Ac expression in the pollen was below the limit of quantitation (LOQ) of the assay used (LOQ = 0.125) (unpublished greenhouse data). It is noteworthy that higher amounts were detected in plant parts preferably attacked by the primary target pest, EFSB. The level of expression of Cry1Ac in Bt eggplant lines tested varied between 0.75±0.33 to 24.87±0.56 ppm DW. These findings are consistent with previous studies conducted in the Philippines [ 25 ] and India [ 21 , 28 ] showing that Bt eggplants have higher levels of Cry1Ac protein expressed in the terminal leaves, flowers and fruits than in the stem and roots. Similarly, a number of studies conducted in other countries also reported variability in Cry protein expression in plant parts in other Bt crops including cotton [ 29 – 31 ], corn [ 32 – 34 ] and rice [ 35 ].
Many researchers have also reported variation in Cry1Ac protein expression in Bt cotton during the growth and development of the plant [ 36 – 42 ]. In Bt cotton, Cry1Ac protein levels were generally high at early stages and then declined as the plant grew to maturity [ 31 , 43 ]. In this study, seasonal variation was also detected in the level of Cry1Ac protein expression in the terminal leaves of Bt eggplant lines. However, the amount of Cry1Ac protein expressed varied only up to 1.7-fold throughout the growing season of 120 days required for profitable eggplant production. Contrary to the results in Bt cotton, in this study the level of Cry1Ac protein expression significantly increased from the vegetative stage to the reproductive stage and either slightly declined or increased at the late reproductive stage depending on the trial. It is important to note that the amount of Cry1Ac protein expressed in Bt eggplant OP lines peaked during the fruit-bearing stage and remained high with the average at 20–23 ppm DW as EFSB pest pressure became more severe.
Factors Affecting Variability in Cry1Ac Protein Expression in Bt Eggplant Lines
Data from other crops also suggest that factors inherent to the variety and the environment affect the variability of Cry1Ac expression. These factors include among others, transgene promoter, parental background, and environmental stressors such as high temperature, heavy drought, waterlogging, and insect damage [ 38 , 44 , 45 ]. In this study, variability in Cry1Ac protein expression in the Bt eggplant lines could also be attributed to using the constitutive 35S CaMV promoter in the EE-1 gene construct, as suggested in studies with Bt cotton which used the same promoter. Parental background has also been reported to affect Cry1Ac protein variability in Bt cotton [ 30 , 31 ]. In this study, the parental background (‘Mara’ and ‘DLP’) of the Bt eggplant OP lines may have influenced, but only to a limited extent, the spatio-temporal variability in Cry1Ac expression. Finally, environmental factors could have contributed to the spatio-temporal expression of the Cry1Ac protein in Bt eggplant lines. Results showed that the levels of Cry1Ac detected during the entire growing season during trial 2 were different compared to results from trial 1. Trial 1 was conducted during the off-season eggplant planting, while trial 2 was conducted during the regular dry season planting. During trial 2, there were more eggplant planted, hence the level of EFSB pest pressure was higher during this season resulting in more damage as shown in the field efficacy data. Weather data obtained during the duration of the two trials indicated that the average daily temperature was similar but the amount of rainfall was much higher in trial 1 than trial 2. A previous report [ 46 ] suggested that environmental factors such as temperature and insect damage could influence expression of a Cry protein.
Variation in Cry1Ac Protein Expression and its Effects on Field Efficacy of Bt Eggplant Lines
It has been a key concern for developers of Bt crops whether variation in Cry protein expression may cause variation in control of the target insect pest. A number of studies in Bt cotton showed that concentration of Cry1Ac correlates well with the efficacy against the target insect pests and that, as the amount of Cry1Ac declines when the crop matures, there is a concomitant decrease in % mortality of the target pest, bollworm ( Helicoverpa armigera or Helicoverpa zea ) [ 29 , 37 , 39 , 47 – 49 ]. In this study, the highest concentration of Cry1Ac protein was expressed in the terminal leaves (24.87± 0.56 ppm DW) and remained high as the Bt eggplant crop matured. The field efficacy of Bt eggplant lines, measured as % EFSB-damaged shoots, also remained very high (95.4–100% reduction) during the entire 10 weeks of evaluation. These results suggest that the high level of expression of Cry1Ac protein results in high field efficacy in Bt eggplant lines. The reduced EFSB-damaged shoots indicate that the effective control of EFSB starting at the vegetative stage will help reduce the field population of EFSB during the fruit-bearing stage resulting in much reduced EFSB damage. Among the plant parts, the level of Cry1Ac protein expressed in the fruits (flesh and skin) was intermediate (2.61±0.36–12.52±3.41 ppm DW). Nevertheless, the % EFSB-damaged fruits in Bt eggplants were effectively reduced (94.1–100% control) throughout the reproductive period of the plants. It should be noted that the lowest concentrations of Cry1Ac detected in the shoots (18.32±2.45 ppm DW) and fruits (2.61±0.36 ppm DW) in the Bt eggplant lines were well above the baseline susceptibility benchmark values of L . orbonalis for Cry1Ac previously reported from India. The average moult inhibitory concentration, MIC 95 , from 29 L . orbonalis populations tested for Cry1Ac was 0.059 ppm [ 21 , 28 ]. More recent work reported the baseline limits for MIC 50 = 0.003 to 0.014 ppm and MIC 95 = 0.028 to 0.145 ppm [ 50 ]. The median lethal concentrations reported were LC 50 = 0.020 and 0.042 ppm [ 50 ] and LC 50 = 0.0326 to 0.0369 mg/mL and LC 90 = 0.0458 to 0.0483 mg/mL of diet [ 51 ]. MIC values have been used in corn as the best estimator of "functional mortality" and predictor of potential effectiveness of Bt corn [ 52 ].
Field Efficacy of Bt Eggplant Lines Containing Event EE-1 Against the Primary Target Pest, ESFB
Efficacy is the capacity of the host plant to affect the survival of the insect pest. Host plant resistance can be measured as a percentage of damage to the foliage or fruiting parts, reduced crop stand, yield and vigor [ 53 ]. It can also be measured based on insect characteristics which include number of eggs laid, aggregation, food preference, growth rate, food utilization, mortality and longevity. In this study, the field efficacy of Bt eggplants against EFSB was evaluated based on the following parameters: (1) percentage of EFSB-damaged shoots; (2) percentage of EFSB-damaged fruits; (3) EFSB larval counts; and (4) survival and fecundity of field collected larvae.
The results of the three season trials indicated consistent, high field efficacy in all Bt eggplant lines tested relative to their non-Bt eggplant comparators i.e. non- Bt near-isoline counterparts and check variety. Even under the most severe pest pressure during trial 2, the Bt eggplant lines demonstrated high level of control of EFSB shoot damage (98.6–100%) and fruit damage (98.1–99.7%) and reduced EFSB larvae infestation (95.8–99.3%). Among the lines tested, Bt M4 showed the lowest % efficacy in shoot (95.3%) and fruit damage (94.1%) and M8 the lowest % efficacy for EFSB larval count (88.4%). However, these lower results were not consistently observed in every trial and their efficacy levels were always much better compared to any of the non-Bt comparators tested.
In addition to Cry1Ac expression and plant damage, we assessed the effect of Bt eggplants on EFSB survivorship and fecundity. This was done to assess the potential for evolution of resistance of EFSB to Bt eggplant. Resistance among insects occur when genetic variation in a population enables a subset of individuals to survive on doses lethal to the majority of the population when feeding on the Bt plant and subsequently produce viable offspring [ 54 , 55 ]. It is noteworthy that results showed few adults emerged and no eggs and viable offspring were produced in mating adults from larvae collected in Bt eggplant lines lending further evidence of very high field efficacy against EFSB. Furthermore, the diminished capacity for normal insect development and reproduction suggest the Bt eggplant lines tested in these trials express a high dose, a key component in the high dose-refuge management strategy [ 56 ].
Taken together, the results obtained from the two-year field testing in Pangasinan support the conclusion that Bt eggplant OP lines developed by the University of the Philippines Los Baños and containing event EE-1 possess a novel trait that provides outstanding control of EFSB making them superior to the conventional counterparts and the check, particularly when the pest pressure is high. Commercial production of Bt eggplant has great potential to reduce yield losses to EFSB while dramatically reducing the reliance of growers on synthetic insecticides to control this pest, reducing risks to the environment, to worker's health, and to the consumer [ 7 , 8 , 10 , 57 ].
Before Bt eggplant seeds are made available for commercial propagation, it is essential to develop an insect resistance management (IRM) plan to manage the risk of resistance evolution in the target pest. The use of high-dose/refuge strategy has been postulated to delay the potential evolution of insect resistance to the Bt crops by maintaining insect susceptibility [ 56 ]. This has been implemented for Bt cotton and Bt corn and the same needs to be extended to Bt eggplants. Some of the key elements in an IRM strategy include information on the expression profile of an insecticidal protein in the Bt crop, the inherent susceptibility of the insect, the number and dominance of genes involved, and the availability of susceptible plants as refuge. Results of the studies presented in this paper indicate that Bt eggplant OP lines expressed the Cry1Ac protein in relevant plant parts primarily attacked by EFSB at the appropriate growth stages throughout the productive life of the crop. More importantly, the amount of Cry1Ac detected in the Bt eggplant shoots and fruits remained sufficiently high to have significant activity against EFSB when compared to the baseline limits previously reported [ 21 , 28 , 50 , 51 ]. Furthermore, the Bt eggplant OP lines exhibited very high levels of field efficacy against EFSB and severely diminished the capacity of EFSB to reproduce successfully.
Prior to the commercial production of Bt eggplant in the Philippines, a structured refuge management strategy will be required. In addition to a structured refuge, the presence of many conventional non-Bt eggplant varieties and alternate wild Solanum hosts commonly present in uncultivated peripheral lands (i.e., unstructured refuges) will serve as a source of susceptible EFSB alleles in the population to slow the evolution of resistance in EFSB. Collectively, the results of this study suggest the possibility of a high-dose/refuge strategy for Bt eggplants. A stringent implementation of high-dose/refuge IRM plan within the context of integrated pest management (IPM) could help delay the potential development of resistance of EFSB to the Bt protein in UPLB Bt eggplant lines.
Materials and Methods
Confined field trials were conducted in the Philippines to evaluate product performance and assess potential environmental risks of UPLB Bt eggplants compared to their non-Bt comparators i.e. non-Bt near-isoline counterparts and the commercial check or reference variety. The field testing site located in the province of Pangasinan, the Philippines, best represented the agro-climatic conditions and production practices in the largest eggplant growing region (Region I or the Ilocos Region) in the country. Pangasinan has Type 1 climate characterized by two pronounced growing seasons: dry, from November to April; wet, during the rest of the year. Eggplant cultivation in Pangasinan is higher during the dry season (DS). Farmers in Pangasinan plant eggplant after rice starting in the months of September to October (planting season) and harvest during the months of December to April. Some farmers also plant during the off-season, which starts at the end of the dry season and harvest during the early wet-season. The province of Pangasinan alone has the widest production area (18.4%) and contributes the largest volume (31.9%) of eggplant produced in the country (2005–2014) [ 5 ]. The Pangasinan field trial site represented the conditions in small-holder farmer’s fields that experience very high natural incidence of EFSB pressure compared with other trial sites.
Three replicated confined field experiments were conducted in Bgy. Paitan, Sta. Maria, for three seasons from March 2010- October 2012. These trials were conducted under natural field infestation of EFSB and without application of lepidopteran-specific insecticide sprays. The studies were conducted in a comparative manner. Bt eggplant lines were evaluated in comparison with the conventional non-Bt comparators consisting of the corresponding non-Bt counterparts with similar genetic backgrounds (recurrent parents/near-isolines) and a National Seed Industry Council (NSIC)-approved commercial open-pollinated variety (OPV) as check or reference genotype. OPVs are standard varieties, which have stable characteristics and produce seeds that will grow into plants more or less identical to their parent plants.
Plant Materials, Experimental Design and Regulatory Conditions
The experimental materials used in the series of three confined field experiments are listed in Table 4 .
1 unsprayed = no lepidopteran-specific insecticide applied
2 BC n = number of backcrossing; F n = filial generation
3 From sowing to end of fallow period
4 D2, D3 = promising advanced Bt OP lines developed through conventional backcross breeding between improved line selection from Dumaguete Long Purple (DLP) and Mahyco eggplant event EE-1; M1, M4, M8 = promising advanced Bt OP lines developed through conventional backcross breeding between improved line selection from cultivar Mara and Mahyco eggplant event EE-1
5 DLP = open-pollinated improved line selection from Dumaguete Long Purple public variety; Mara, Mara S1, Mara S2 = open-pollinated improved line selections from the Mara cultivar developed by UPLB-IPB Vegetable Breeding Division
6 National Seed Industry Council (NSIC)-registered commercial open-pollinated eggplant variety
Plant materials
The Bt eggplant OP lines (D2, D3, M1, M4, M8) used as test entries in the field trials are advanced breeding lines (BC 3 F 4 to BC 3 F 6 ) derived from initial crosses of Mara selection x Mahyco elite line, ‘EE-1’ and DLP selection x Mahyco elite line, ‘EE-1’. The non-Bt comparators were: (1) DLP as the non-Bt counterpart genotype of Bt D2 and D3 OP lines; (2) Mara, Mara 1 or Mara S2 as the counterpart genotypes for M1, M4 and M8; and (3) Mamburao, a non-Bt eggplant OP variety approved by the NSIC [ 58 ] as the check or reference genotype.
Experimental Design and Field Layout
Each field experiment was planted in randomized complete block design (RCBD) with four replications, 4–6 rows/plot and 10 plants per row. The perimeters of each field experiment were surrounded by five rows (1 m between rows) of conventional non-Bt eggplant OP as pollen-trap plants. The experimental set up was conducted in a fenced facility with restricted access. A 200-meter radial distance isolated the field trial site from the nearest eggplants in the area.
Permissions
All field trials were conducted in accordance with the Department of Agriculture Administrative Order No. 8 Series of 2002 for field testing ( www.biotech.da.gov.ph ). The Bureau of Plant Industry (BPI) issued the corresponding Biosafety Permit for Field Testing in Bgy. Paitan, Sta. Maria, Pangasinan. The biosafety permit conditions were complied with throughout the conduct of every field experiment and associated greenhouse and laboratory activities. All sample collection and transport of materials were done under the supervision of the duly designated biosafety trial inspectors following the prescribed biosafety procedure for sample collection, handling and transport.
Crop Establishment, Management, Harvesting and Termination
Seedling establishment.
Seeds of UPLB Bt and non-Bt eggplant entries (treatments) were sown in pots with sterilized soil 30–34 days before transplanting. The germinated seeds were pricked (transferred individually in seedling trays), 7–8 days after sowing (DAS) and maintained inside the BL2 greenhouse at UP Los Baños. At 28–30 DAS, representative seedlings for the seed lot of each entry were tested for presence or absence of Cry1Ac using immunoassay or gene strip test kit, DesiGen Xpresstrip (DesiGen, Maharashtra, India), as described in Ripalda et al . [ 25 ]. Excess transgenic seedlings were disposed of properly in a disposal site inside the BL2 greenhouse. Seedlings of Bt and non-Bt eggplant test entries, check varieties and pollen traps were transported from UP Los Baños to the confined field testing site in Bgy. Paitan, Sta. Maria, Pangasinan for transplanting.
Cultural management
The confined field trials were managed based on the national cooperative trial guidelines for eggplant [ 59 ] and prevalent agronomic practices for eggplant growing in the region, including site preparation, tillage, and nutrient applications. Manual watering of plants was done during the first month after transplanting and shifted to overhead and/or furrow irrigation as plants grew and required greater amounts of water. At times of continuous heavy rain during the trial period, trenches or canals were dug to keep the soil near the roots from being waterlogged and to reduce the incidence of bacterial wilt infection. Staking of plants was done to provide additional support as the number and size of fruits increase and during periods of strong winds and rain. Branches were kept off the ground to prevent the fruit from becoming deformed.
Pest management
No lepidopteran-specific insecticide sprays were applied during the entire duration of the trials. Management of other arthropod pests and diseases was done by application of recommended IPM practices, primarily sanitation and withholding of pesticide use as long as possible to enable the proliferation of natural enemies. Whenever populations of leafhoppers and mites rose to very high level, they were controlled with the application of insecticides with reduced risk and without activity against EFSB (a.i. thiamethoxam) and sulphur, respectively.
Termination, disposal and fallow period
After the final harvest, each field experiment was terminated. All above- and below- ground plant parts were removed from the field and disposed of properly following the prescribed procedure indicated in the Biosafety permit. The field was plowed, irrigated and observed for volunteer plants 7, 14 and 30 and 60 days after termination. The field was kept fallow for at least 60 days after termination.
Data Collection and Analysis
Determination of cry1ac protein expression.
Cry1Ac protein expression study was conducted in trial 1 and trial 2, which represented the two eggplant growing seasons in Pangasinan, i.e. wet/off-season and regular dry season planting, respectively. Because trial 3 was also planted during the same season as trial 1 (wet/off-season) and for cost consideration, Cry1Ac protein analysis was not performed from samples obtained in this trial.
Sample collection used was based on the protocol previously described in Ripalda et al . [ 25 ]. Different plant parts from at least five plants from among the 16 plants in the two inner rows/replicate plot were collected. Terminal leaves were collected at the vegetative (up to 25 days after transplanting, DAT), reproductive (25–60 DAT) and late reproductive (60–80 DAT) stages of the crop. Flowers and immature fruits were collected during reproductive and late reproductive stages. Fruit samples were collected during the harvest period. Stem and roots were collected at termination (around 150 DAT). All samples collected were kept in an icebox and transported to the laboratory. Flesh and skin of immature fruits, but avoiding seeds, were separated in thin slices. Stems and roots were washed prior to storage. The woody portion of the stems was used for analysis. All samples were kept in a -80°C biofreezer until further processing and then freeze-dried at -60°C for 1–5 days until crisp. Dried samples from three plants per plot per plant part were bulked and placed in a 2.0 mL microfuge tube. Bulked samples were homogenized using two 6-mm steel beads and the ground samples were put in sealed containers and stored at 4°C until use.
Quantification of Cry1Ac was done through an enzyme-linked immunosorbent assay (ELISA). Commercially available quantitative ELISA kits (DesiGen Cry1Ac QuanT) specific for the Cry1Ac protein were procured from Mahyco (Maharashtra, India). Five milligrams of the powdered samples were weighed and analyzed. Chilled extraction buffer prepared as specified in the kit was added to the weighed samples. A dilute (up to 1:8) trypsinized protein extract was loaded to the pre-coated plates. Positive and negative controls and standards were prepared and loaded according to the instructions in the kit. Antibodies, wash buffer and substrate for detection (pNPP) used were also from the kit. Absorbance readings of the samples were made at 405 nm. According to the manufacturer’s instruction, the assay is considered valid when the mean absorbance reading of the blank is ≤0.246, mean absorbance reading of the standards with the highest concentration of Cry1Ac is ≥1.305, % residual of back calculated concentration of standards are 20 ng/ml– 125 ng/ml standards: ≤15%; 0.625 ng/ml standard ≤25% and R 2 of the standard curve is ≥0.98.
Data on Cry1Ac concentrations from different plant parts and different developmental stages of the Bt eggplant lines were analyzed by one-way analysis of variance using PROC MIXED in SAS v.9.1.3 [ 60 ]. Means were separated using Tukey’s HSD at α = 0.05. Data available from the Dryad Digital repository: http://dx.doi.org/10.5061/dryad.ks131 [ 61 ]
Evaluation of Field Damage by EFSB
Fruits were harvested from the 16 plants located in the inner two rows (12 m 2 ) of each plot. During each harvest period, the harvested fruits per plot were carefully cut open and examined for the presence of EFSB larvae or signs of EFSB damage and tunneling, sorted as with or without EFSB-damage, counted and weighed separately.
Data gathered:
- Percentage (%) damaged shoots per plot–calculated from the number of damaged shoots due to EFSB recorded from five shoots per plant from 16 inner row plants per plot at weekly intervals starting at two weeks after transplanting (WAT) for 10 observation periods.
- Percentage (%) damaged fruits per plot—calculated from the total number of EFSB-damaged fruits over the total number of fruits harvested from 16 inner row plants per plot. Harvesting was done every 3–4 days. Data were collected from 10–17 harvest periods prior to the termination of the experiment.
- EFSB larval counts (no. larvae/plot)–All harvested fruits from the 16 inner row plants per were cut open to check for the presence of EFSB larvae. The number of surviving EFSB larvae found inside the fruits per replicate plot were recorded every harvest period.
- % Efficacy (or Level of control)–calculated based on the formula (1- Bt/nonBt)*100% for % EFSB-damaged shoots, damaged fruits and EFSB larval counts
- Survivorship and fecundity–All surviving larvae collected per plot per harvest period were transferred to individual plastic cups and labeled. Each cup was provided with a slice of eggplant fruit from which the larva was collected. The cups were then brought to the UPLB-IPB Entomology P2 Laboratory and reared continuously in their respective hosts (Bt or non-Bt) until the adult stage. The number of individuals that successfully reached pupal and adult stages was recorded. Pairs of surviving adults from Bt and from conventional non-Bt lines were mated, placed in oviposition chambers and observed for egg deposition and hatching of offspring.
Data transformation was used to improve the normality of variables due to markedly skewed data or heterogeneous variances of Bt and conventional non-Bt entries. Data collected were transformed to sqrt (Y+0.5), arcsin (sqrt(Y/100)) or log 10 (Y+1) as appropriate. Transformed data on percentages damaged shoots and fruits, larval counts and feeding tunnel lengths were subjected to one-way analysis of variance and analyzed using PROC MIXED in SAS v.9.1.3 [ 60 ] and means were separated using Tukey’s HSD at α = 0.05. Pairwise mean comparisons by contrast between each Bt and its respective non-Bt counterpart and check variety were done for all parameters gathered using PROC MIXED. Data available from the Dryad Digital repository: http://dx.doi.org/10.5061/dryad.ks131 [ 61 ]
Supporting Information
Trials 1 to 2. CY 2010–11, Sta Maria, Pangasinan, Philippines.
Trials 1 to 3. CY 2010–12, Sta. Maria, Pangasinan, Philippines.
A: Trial 1; B: Trial 2; C: Trial 3. CY 2010–12, Sta. Maria, Pangasinan, Philippines.
Acknowledgments
We gratefully acknowledge the critical support and valuable contributions of these institutions. This research was co-funded by the United States Agency for International Development (USAID) through Cornell University Agricultural Biotechnology Support Project II (ABSPII), the Republic of the Philippines Department of Agriculture- Biotechnology Program Office (DA- Biotech BPO) and the Instiute of the Plant Breeding, College of Agriculture, University of the Philippine Los Baños (UPLB). We thank the Maharashtra Hybrid Seeds Co. Pvt. Ltd. (Mahyco) for providing access to eggplant event EE-1 and regulatory-related information, and for various technical assistance/advice in the conduct of laboratory and field activities; and Cornell University and Sathguru Management Consultants for facilitating the technology transfer. We also acknowledge the assistance of UPLB Foundation Inc., the executing agency for the ABSPII project in the Philippines.
We sincerely thank the following Bt eggplant staff for their various valuable contributions: OO Silvestre, MMM Abustan, RN Candano, RP Urriza, ER Maligalig, SA Baldo and MM Marin for field establishment and management and lab/field data collection; RR Ripalda, RB Frankie, ML de Vera and AD Austral for sampling and Cry1Ac analysis; and ZJ Bugnosen for assistance in regulatory-related and administrative activities. We thank B. Bartolome for statistical advice and JA Estrella and RN Candano for assistance in statistical analyses; and N. Storer for reviewing an earlier draft of the manuscript. Special thanks to the people of Sta. Maria, Pangasinan especially Ret. Gen. M Blando, Dr. R Segui for all forms of assistance during the conduct of the trials.
Funding Statement
This research was funded through the United States Agency for International Development (USAID Cooperative Agreement GDG-A-00-02-00017-00) to Cornell University Agricultural Biotechnology Support Project II (ABSPII), and matching funds from the Republic of the Philippines Department of Agriculture- Biotechnology Program Office (DA- Biotech BPO) and the University of the Philippine Los Banos (UPLB). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability
Click through the PLOS taxonomy to find articles in your field.
For more information about PLOS Subject Areas, click here .
Loading metrics
Open Access
Peer-reviewed
Research Article
Bt eggplant ( Solanum melongena L.) in Bangladesh: Fruit production and control of eggplant fruit and shoot borer ( Leucinodes orbonalis Guenee), effects on non-target arthropods and economic returns
Roles Formal analysis, Investigation, Writing – original draft
Affiliation Tuber Crops Research Sub Centre, BARI, Bogra, Bangladesh
Roles Data curation, Investigation
Affiliation On Farm Research Division, BARI, Bogra, Bangladesh
Roles Investigation
Roles Project administration
Affiliation BARI, Joydebpur, Gazipur, Bangladesh
Affiliation Country Coordinator for Bangladesh, USAID Feed the Future South Asia Eggplant Improvement Partnership, Dhaka, Bangladesh
Roles Formal analysis, Methodology, Writing – review & editing
Affiliation USDA-ARS, Arid-Land Agricultural Research Center, Maricopa, Arizona, United States of America
Roles Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Writing – review & editing
* E-mail: [email protected]
Affiliation Department of Entomology, Cornell University/New York State Agricultural Experiment Station (NYSAES), Geneva, New York, United States of America
- M. Z. H. Prodhan,
- M. T. Hasan,
- M. M. I. Chowdhury,
- M. S. Alam,
- M. L. Rahman,
- A. K. Azad,
- M. J. Hossain,
- Steven E. Naranjo,
- Anthony M. Shelton
- Published: November 21, 2018
- https://doi.org/10.1371/journal.pone.0205713
- Reader Comments
Eggplant or brinjal ( Solanum melongena ) is a popular vegetable grown throughout Asia where it is attacked by brinjal fruit and shoot borer (BFSB) ( Leucinodes orbonalis ). Yield losses in Bangladesh have been reported up to 86% and farmers rely primarily on frequent insecticide applications to reduce injury. Bangladesh has developed and released four brinjal varieties producing Cry1Ac (Bt brinjal) and is the first country to do so. We report on the first replicated field trials comparing four Bt brinjal varieties to their non-Bt isolines, with and without standard insecticide spray regimes. Results of the two-year study (2016–17) indicated Bt varieties had increased fruit production and minimal BFSB fruit infestation compared with their respective non-Bt isolines. Fruit infestation for Bt varieties varied from 0–2.27% in 2016, 0% in 2017, and was not significantly affected by the spray regime in either year. In contrast, fruit infestation in non-Bt lines reached 36.70% in 2016 and 45.51% in 2017, even with weekly spraying. An economic analysis revealed that all Bt lines had higher gross returns than their non-Bt isolines. The non-sprayed non-Bt isolines resulted in negative returns in most cases. Maximum fruit yield was obtained from sprayed plots compared to non-sprayed plots, indicating that other insects such as whiteflies, thrips and mites can reduce plant vigor and subsequent fruit weight. Statistically similar densities of non-target arthropods, including beneficial arthropods, were generally observed in both Bt and non-Bt varieties. An additional trial that focused on a single Bt variety and its isoline provided similar results on infestation levels, with and without sprays, and similarly demonstrated higher gross returns and no significant effects on non-target arthropods. Together, these studies indicate that the four Bt brinjal lines are extremely effective at controlling BFSB in Bangladesh without affecting other arthropods, and provide greater economic returns than their non-Bt isolines.
Citation: Prodhan MZH, Hasan MT, Chowdhury MMI, Alam MS, Rahman ML, Azad AK, et al. (2018) Bt eggplant ( Solanum melongena L.) in Bangladesh: Fruit production and control of eggplant fruit and shoot borer ( Leucinodes orbonalis Guenee), effects on non-target arthropods and economic returns. PLoS ONE 13(11): e0205713. https://doi.org/10.1371/journal.pone.0205713
Editor: Juan Luis Jurat-Fuentes, University of Tennessee, UNITED STATES
Received: May 8, 2018; Accepted: October 1, 2018; Published: November 21, 2018
Copyright: © 2018 Prodhan et al. This is an open access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability: The authors from the Bangladesh Agricultural Research Institute (BARI) collected and analyzed the data. Advice on the experimental design and data analysis were provided by Drs. Shelton and Naranjo who also helped write the paper. The processed data can be obtained at doi: 10.5061/dryad.q4b9r2k .
Funding: The authors gratefully acknowledge the support provided by the United States Agency for International Development (USAID) for their Feed the Future South Asia Eggplant Improvement Partnership (AID-OAA-A-15-00052). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Genetically engineered (GE) crops continue to expand and transform agriculture on a global scale. In 2017, nearly 190M hectares of GE crops were grown by about 18M farmers in 24 countries, including 101M hectares of crops with high levels of host plant resistance to caterpillar and beetle pests [ 1 ]. Between 1996 and 2015, this adoption has been associated with increases in farm income > $50,274M and $45,958M, in Bt cotton and maize, respectively, and reductions of > 268M and 87M kg of insecticide active ingredient in Bt cotton and maize, respectively [ 2 ]. However, the potential benefits provided by Bt crops have largely gone unrealized in fruits and vegetables where insect management continues to rely primarily on the use of synthetic insecticides [ 3 ]. This situation is changing in Bangladesh with the introduction of Bt eggplant.
Eggplant ( Solanum melongena L.), or brinjal as it is called in Bangladesh and India, is one of the most important and popular vegetables in South and Southeast Asia. The crop is damaged severely by the brinjal fruit and shoot borer (BFSB)( Leucinodes orbonalis Guenee) (Lepidoptera: Crambidae). The caterpillar damages brinjal by boring into the petiole and midrib of leaves and tender shoots, resulting in wilting and desiccation of stems. Larvae also feed on flowers, resulting in flower drop or misshapen fruits. The most serious economic damage caused by BFSB is to the fruit, because the holes, feeding tunnels, and larval excrement may make the fruit unmarketable and unfit for human consumption.
BFSB poses a serious problem because of its high reproductive potential, rapid turnover of generations and intensive damage during the wet and dry seasons. Infestation levels may exceed 90% and the yield loss has been estimated up to 86% in Bangladesh [ 4 ]. It has been reported that 98% of Bangladeshi farmers rely solely on insecticide sprays to control BFSB [ 5 ] and farmers may apply as many as 84 insecticide sprays during the cropping season [ 6 ]. This practice is unhealthy for consumers, farmers, and the environment, and is expensive to farmers. As an alternative to intensive use of insecticides, the India-based Maharashtra Hybrid Seed Company (Mahyco) inserted the Cry1Ac gene, under the control of the constitutive 35S CaMV promoter, into eggplant (termed ‘event’ EE-1) to control feeding damage by EFSB. Bt eggplant demonstrated control of EFSB in contained greenhouse trials in India [ 7 ]. In late 2003, a partnership was formed between Mahyco, Cornell University, United States Agency for International Development (USAID), and public sector partners in India, Bangladesh, and the Philippines under the Agricultural Biotechnology Support Project II [ 7 ]. Mahyco donated the EE-1 event to the Bangladesh Agricultural Research Institute (BARI), where it was incorporated into BARI-developed local varieties. Subsequently, BARI applied to the National Technical Committee on Crop Biotechnology (NTCCB) to release Bt eggplant. Following the recommendation from the NTCCB, the application for release was forwarded to the National Technical Committee on Crop Biotechnology (NTCCB) Core Committee followed by the National Committee on BioSafety (NCB). The Bangladesh government granted approval for release of four varieties on 30 October 2013. On 22 January 2014, Bt seedlings were distributed to 20 farmers in four districts making Bangladesh a pioneer in the world to allow the commercial cultivation of a genetically engineered vegetable crop.
The effective use of this technology requires important knowledge of the agronomic nature of the four varieties and their ability to control BFSB. Furthermore, information is needed on how Bt brinjal affects non-target pest arthropods that likely can affect the yield and quality of brinjal. Performance of the four Bt brinjal varieties and their isolines in spray and no-spray conditions should provide information about the potential damage by these other pests. Likewise, it is important to assess the effect of Bt brinjal on beneficial arthropods, especially biological control agents that can help suppress pest populations. Here we report on the first replicated field trials in Bangladesh to assess the ability of the four Bt brinjal varieties to control BFSB, with and without a standard insecticide regime, compared to their non-Bt isolines. In addition, we assessed plant growth characteristics, economic returns, and potential effects on non-target arthropod pests and on beneficial arthropods that might supply important biological control services.
Materials and methods
Two sets of complementary experiments were conducted over a two-year period (2016–7) in Bangladesh. In the first experiment, the four commercialized Bt lines were compared to their non-Bt isolines, with and without insecticide sprays to: a) assess their ability to protect the plant from EFSB, b) assess their agronomic characteristics, c) document effects on other arthropods, and d) assess their economic return. In the second experiment, a single Bt line was compared to its isoline, with and without insecticide sprays. This experiment placed more emphasis on assessing the effects of the lines and spray treatments on non-target arthropods, while also assessing the ability of the treatments to control EFSB and provide favorable economic returns.
Comparisons of four Bt Brinjal varieties and their isolines
Plants, sprays and experimental design..
Experiments were conducted at the On-Farm Research Division (OFRD) of BARI, in the Bogra District (089 0 22.922 l E; 24 0 51.056 l N) of Bangladesh. In both years, the experimental field was laid out in a randomized complete block split-plot design with four replications that included insecticide spray regimes as main plots and varieties as sub-plots.
The trials utilized the four Bt brinjal varieties released by BARI to farmers in 2014 [ 7 ]: BARI Bt begun-1, BARI Bt begun-2, BARI Bt begun-3, BARI Bt begun-4) and their respective non-Bt isolines (BARI begun-1, BARI begun-2, BARI begun-3, BARI begun-4). In each year, the experimental area was ca. 0.1 ha. Main plots receiving insecticides treatments were 6.0 m x 12.0 m with 3.0 m × 3.0 m sub-plots receiving the brinjal varieties. The distances between sub-plots, main-plots and blocks were 30.0 cm, 1.0 m and 1.5 m, respectively. Row-to-row and plant-to-plant spacing was 100 cm and 75 cm, respectively. Seedlings 35-days old) were transplanted on 12 January 2016 and 10 February 2017.
Sprayed plots were treated weekly with both Admire 20SL (imidacloprid) at 0.5 ml/L of water (active ingredient 50 ml/ha) for sucking arthropods (whiteflies, mites, jassids and aphids) and Proclaim 5SG (emamectin benzoate) at 1 g/L of water (active ingredient 25 g/ha) for BFSB. These two insecticides are commonly used in brinjal production in Bangladesh and weekly, or more frequent, spray schedules are the norm. Sprays were applied using a Knapsack sprayer. Spraying started from crop establishment and continued at weekly intervals to the last harvest, 30 May in 2016 and 25 June in 2017. Before spraying, the non-sprayed plots were covered with a non-porous cloth to prevent spray drift. Non-sprayed plots were sprayed with water only. Fertilizers were used at 138-40-100-18-1.7–3.6 kg/ha (NPKSBZn) and cowdung at10 t/ha. Irrigation, weeding, pruning of side shoots and other cultural operations were done when necessary following standard practices for brinjal production in Bangladesh [ 8 ].
Measurements and data analysis.
Data were collected weekly on plant growth patterns (plant height and width (or bushiness)), number of flowers per plant, percent damaged shoots and fruits by BFSB, and marketable and non-marketable fruits on each of four plants per plot. Arthropod populations (pests and beneficials) were sampled weekly on the five newest leaves on each of four randomly selected plants per plot. The upper and lower surfaces of the leaves were thoroughly examined for the presence of arthropods. All weekly counts were taken from 1 February to 30 May, 2016 and from 1 April to 25 June, 2017. Arthropod counts were made before 9 am. A mixed-model, split-plot ANOVA was used for analyses with block as the random effect and plant type and insecticide as fixed effects. Each year was analyzed separately. The response variable was the seasonal mean for each variable examined over time. Arcsine square-root transformations were applied to percentage data but untransformed means are presented. Mean differences were contrasted using Tukey’s HSD test and analyses were done using the statistical package ‘R’. The total seasonal pesticide load (each insecticide applied × number of applications × dose) was used to estimate the Environmental Impact Quotient (EIQ) [ 9 ].
Comparisons of one Bt Brinjal variety and its isoline
The experiment was conducted at OFRD, BARI, Bogra (089 0 22.858 ′ E; 24 0 51.088 ′ N), Bangladesh. In both years, the experiment was laid out in a ca. 0.1 ha field using a randomized complete block design with four replications. Plot size was 4.5m × 9.0 m and the distances between plots and blocks were 30.0 cm and 1.5 m, respectively. Treatments consisted of two varieties of brinjal, BARI Btbegun-1 and BARI begun-1 (non-Bt isoline), each sprayed or unsprayed with insecticides for a total of four treatments. Seedlings were transplanted on January 13, 2016 and on 11 February 2017. Plant spacing, fertilizer use, cultural practices and insecticide sprays were as described above.
Data were taken weekly on percent damaged shoots and fruits by BFSB, marketable and non-marketable fruits, economic returns and densities of arthropods (pest and beneficial arthropods). Arthropod populations were assessed using three methods: 1) Counts on plants were taken from 10 randomly selected plants from the interior of the plot. For each plant, all arthropods were counted on the upper and lower surface of the top 5 leaves and counts were made before 9 am; 2) yellow sticky traps (45×18 cm) were used to measure aerial populations of insects with three traps placed in each plot at crop canopy level; 3) pitfall traps were used to measure ground dwelling arthropods. Three plastic cups (10 cm diameter and 8 cm deep) were placed in the soil in each of the plots with the mouth of the cup at ground level. Each cup was half-filled with water and a few drops of detergent as a trapping fluid. Each week the trapped arthropods were placed into plastic bottles filled with 70% alcohol. Samples were labeled and stored until identified. All counts were done weekly from 2 February to 31 May, 2016 and from 1 April to 25 June, 2017. A mixed-model ANOVA was used with block as the random effect and plant type and insecticides as fixed effects. Each year was analyzed separately. The response variable was the seasonal mean for each variable examined over time. Arcsine square-root transformations were applied to percentage data but untransformed means are presented. Mean differences were contrasted using Tukey’s HSD test and analysis were done using the statistical package ‘R’. The total seasonal pesticide load (each insecticide applied × number of applications × dose) was used to estimate the Environmental Impact Quotient (EIQ) [ 9 ]. As before, gross returns and gross margins were estimating with a partial budgeting analysis.
Infestation by BFSB.
In both years, significant differences were observed among the varieties for BFSB infestation ( Table 1 ). Regardless of spray regime, there was no shoot infestation by EFSB in any of the four Bt brinjal varieties in either year, but shoot infestation occurred in all non-Bt brinjal varieties regardless of spray regime. Infested fruit for Bt varieties varied from 0 to 2.27% and was not significantly affected by the spray regime in either year. In contrast, the percent infested fruit in the non-Bt brinjal varieties reached 36.70% in 2016 and 45.51% in 2017 ( Table 1 ). Another measure of fruit infestation was the percent fruit infested by weight, which was more reflective of income because brinjal is sold by weight and infested fruit bring a lower value. By weight, the highest percent of infested Bt brinjal fruit was only 2.27% (2016, BARI Bt begun 2), compared to the highest rate for non-Bt brinjal of 44.30% (2017, non-Bt isoline 1). In some cases, spraying significantly reduced infestation in non-Bt varieties [e.g. 2016, sprayed non-Bt isoline 4 reduced damage by 25.38% (34.49–9.11%) while in other cases spraying did not (e.g. 2016, non-Bt isoline 2 (21.89–19.17%)].
- PPT PowerPoint slide
- PNG larger image
- TIFF original image
https://doi.org/10.1371/journal.pone.0205713.t001
Yields and gross margins.
In both years, significant differences were evident in the economic returns due to BFSB infestation and costs for spraying ( Table 2 ). In both years all Bt lines has higher gross margins than their isolines, regardless of whether they were sprayed or not. In 2016, all four Bt brinjal varieties showed a positive gross margin, even when no sprays were applied. In contrast only two of the non-Bt isolines that were sprayed showed a positive gross margin when sprayed and only one of the unsprayed non-Bt isolines showed a positive gross margin. In 2017, all of the non-sprayed, non- Bt isolines had negative gross margins, as did one of the non-sprayed Bt brinjal varieties (BARI Bt begun-4). In 2016, spraying Bt brinjal varieties always improved the gross margin, despite the higher production costs. Spraying a non-Bt isoline could also improve its gross margin, but not to the level of the sprayed Bt variety (e.g., in 2016 the gross margin for sprayed Bt begun-2 was $7,634.74 compared to its sprayed non-Bt isoline of $2,458.00).
https://doi.org/10.1371/journal.pone.0205713.t002
Effects on non-target pest arthropods.
Eleven different non-target pest arthropods were observed. Five sucking pests, including whitefly ( Bemisia tabaci Gennadius), thrips ( Thrips palmi Karny), aphid ( Aphis gossypii Glover), jassid ( Amrasca biguttula biguttula Ishida) and mites ( Tetranychus urticae Koch) had populations > 0.2 per leaf per and were analyzed ( Table 3 ). Populations of flea beetle ( Phyllotreta striolata ), armyworms ( Spodoptera litura ), Mirid bug ( Helopeltis sp.), Epilachna beetle ( Epilachna sp.) and stink bug ( Nezara sp.) were too low (< 0.01) for meaningful analysis. In 2016, spraying generally significantly increased populations of whiteflies and mites but decreased populations of thrips, aphids and jassids. While there were significant differences in other non-target pest populations, they were always at low densities and effects were likely of little consequence. In 2017, generally there were statistically higher populations of whiteflies, but statistically lower populations of aphids and jassids in sprayed plots. Mites were not present in 2017 probably because of the wetter weather. Variety did not have a consistent effect on densities of non-target pest arthropods.
https://doi.org/10.1371/journal.pone.0205713.t003
Effects on non-target beneficial arthropods.
Ladybird beetles and spiders were the most abundant beneficial arthropods but still only reached 0.067 beetles/leaf and 0.030 spiders/leaf in 2016 ( Table 4 ). Populations of red ant ( Solenopsis sp.), rove beetle ( Homaeotarsus sp.), assassin bug ( Zelus sp.), ground beetle ( Ophionia nigrofasciata ,), syrphid fly ( Syrphus sp.) and small black ant ( Camponotus sp.) were too low (< 0.01) for meaningful analysis. In 2016 spraying sometimes, but not consistently, significantly reduced the populations of Coccinella sp. but not spiders. In 2017, there were no significant differences between populations of Coccinella and spiders on a variety in sprayed and non-sprayed plots. Variety did not have a consistent effect on non-target beneficial arthropods.
https://doi.org/10.1371/journal.pone.0205713.t004
Variety characteristics.
There were some significant differences in plant height, bushiness (width) and number of shoots and flowers per plant between Bt varieties and their non-Bt isolines ( Table 5 ). However, there were no clear trends that Bt plants differed from their isolines in height or bushiness, although it has been suggested that BFSB-infested shoots may affect plant architecture by killing stems. Likewise, spraying did not appear to have a consistent effect on plant characteristics. The most dramatic and consistent differences were the number of fruit per plant. Generally, Bt plants had significantly more fruit per plant than their respective non-Bt isolines in both years.
https://doi.org/10.1371/journal.pone.0205713.t005
Environmental impact quotient.
In both years, the same number of sprays (19) was applied to sprayed plots. The seasonal insecticide load/ha for imidacloprid and emamectin benzoate was 36.7 and 26.3 mg active ingredient per ha, respectively, and the seasonal calculated EIQ values for imidacloprid were 8.4 (consumer), 5.6 (field worker) and 75.5 (ecological) and for emamectin benzoate 1.7 (consumer), 3.8 (farmer worker) and 27.9 (ecological).
Comparisons of one Bt Brinjal varieties and its isoline
In both years, in all but one case (i.e., sprayed plots in 2016) there were significant differences in infested shoots and fruit for Bt begun-1 compared to its isoline ( Table 6 ). In 2016, percent infested fruit for Bt begun-1varied between 0–0.16% depending on whether it was sprayed or not, while its non-Bt isoline had infestation rates between 39.33 and 50.85% when sprayed or not, respectively. In 2017, similar lack of infestation of Bt begun-1 fruit was observed whether it was sprayed or not, while its isoline had 41.52% infested fruit when sprayed and 52.43% when not sprayed ( Table 6 ).
https://doi.org/10.1371/journal.pone.0205713.t006
In 2016 Bt begun-1 had a higher gross margin than its isoline. Spraying Bt begun-1 improved the gross margin to $2,962.94 /ha compared to not spraying Bt begun-1 ($939.42/ha), despite the increased cost of production ( Table 7 ). Likewise, in 2017 spraying Bt begun-1 improved the gross margin from $1,289.78 to $3,654.59. In both years, not spraying the non-Bt line resulted in a negative gross margin.
https://doi.org/10.1371/journal.pone.0205713.t007
Eleven non-target pest arthropods species were observed on leaves in 2016, with whiteflies, aphids, thrips, jassids, flea beetles and mites having sufficient populations for meaningful analysis ( Table 8 ). Only with jassids and mites did variety have a significant effect, but it was not consistent. Spraying increased whiteflies abundance in both years, but this was not the case with most other species.
https://doi.org/10.1371/journal.pone.0205713.t008
Populations of mirid bug ( Helopeltis sp.), monolepta beetles ( Monolepta sp), shield bug ( Scutiphora sp.), leaf miner ( Liriomyza sp.) and green leafhopper ( Nephotettix sp.) were too low (< 0.01) for meaningful analysis. In 2017, populations of flea beetle ( Phyllotreta striolata ), Mirid bug ( Helopeltis sp.), Epilachna ( Epilachna sp.), Bombardier beetle ( Pheropsophus sp.), Hooded hopper ( Oxyrachis terandus Fab.), Semiloper ( Trichoplusia sp.), mites ( Tetranychus urticae Koch) and green leaf hopper ( Nephotettix sp.) were too low (< 0.01) for meaningful analysis.
In 2016, there were only six non-target species of pest arthropods caught in pitfall traps: flea beetles, grasshoppers, monolepta beetles, termites, June beetles and stink bugs, and their populations were all too low for meaningful analysis. Sticky traps captured aphids, whiteflies, flea beetles and jassids but populations were too low for meaningful analysis. Similar results were observed in 2017 with all different sampling methods.
As in the first experiment, lady beetles and spiders were the most abundant beneficial arthropods in 2016 on plant samples, but only reached a peak of 0.04 beetles/leaf and 0.029 spiders/leaf ( Table 9 ). Spraying did not have a consistent effect on reducing the densities of either beneficial in either year. Populations of red ant ( Solenopsis sp.), small black ant ( Camponotus sp.), rove beetle ( Homaeotarsus sp.), ground beetle ( Ophionia nigrofasciata sp.) and syrphid fly ( Syrphus sp.), honeybee ( Apis sp.) and bombardier beetle ( Pheropsophus sp.) were too low (<0.01) for meaningful analysis. For pitfall traps, the numbers of beneficial arthropods captured were too low (<0.1/trap) for meaningful analysis.
https://doi.org/10.1371/journal.pone.0205713.t009
Yellow sticky traps caught flying insects in the crop canopy but the numbers were low, with the highest counts being ladybird beetles at 0.446 per trap/wk. ( Table 10 ). In neither year were populations affected by insecticide sprays or brinjal variety. Populations of rove beetle ( Homaeotarsus sp.), damsel fly ( Agriocnemis sp.) and ground beetle ( Ophionia nigrofasciata ) were too low (<0.01) for meaningful analysis.
https://doi.org/10.1371/journal.pone.0205713.t010
Environmental impact quotient (EIQ).
The same number of sprays (19) was applied to sprayed plots in both years. The seasonal insecticide load/ha for imidacloprid and emamectin benzoate was 36.7 and 26.3 mg active ingredient per ha, respectively, and the seasonal calculated EIQ values for imidacloprid were 8.4 (consumer), 5.6 (field worker) and 75.5 (ecological) and for emamectin benzoate 1.7 (consumer), 3.8 (farmer worker) and 27.9 (ecological). This was the same as in the first experiment with four varieties.
These studies present the first replicated field trials assessing the four Bt brinjal varieties that were first introduced to Bangladesh farmers in 2014. Collectively, the results demonstrate that the four Bt varieties provided more fruit and nearly complete protection from infestation by BFSB, compared to their non-Bt isolines, even when no insecticide sprays were applied. Most importantly, these studies revealed that all Bt lines had higher gross returns than their non-Bt isolines.
The insecticide spray regime of using imidacloprid and emamectin benzoate on non-Bt brinjal was unable to decrease the level of BFSB infestation to the level achieved by using its Bt brinjal isoline. Furthermore, it is worth noting that spraying these insecticides tended to increase the infestation in non-Bt fruit. For example, in Table 1 for 2016 the average percent of infested fruit by weight of all four non-Bt lines was 28.2% when they were sprayed and only 16.5% when not sprayed. In 2017 the same phenomenon occurred with an average infestation of 38.4% when sprayed and only 18.9% when not sprayed. One hypothesis for this phenomenon is that spraying reduced the natural enemy population of BFSB and thus increased damage to the brinjal. Further work is needed to confirm this hypothesis.
When comparing the arthropod communities in Bt and non-Bt brinjal, we were not able to detect any differences in numbers of either non-target pest species or beneficial species, suggesting that the four Bt brinjal varieties control the most important insect pest of brinjal in Bangladesh, BFSB, without disrupting arthropod biodiversity. However insecticide sprays did have a disruptive effect on some species of beneficial arthropods and this could support the hypothesis proposed above.
It is important to note that the yield of Bt brinjal, based on weight of the fruit, was improved with the insecticide spray regime. It appears that arthropods such as whiteflies, mites, jassids and aphids, none of which are susceptible to Cry1Ac, still need to be managed. Scheduled applications of the two insecticides, without regard to any threshold, resulted in a relatively high EIQ. The next challenge will be to develop thresholds for the common sucking arthropods encountered in Bangladesh using selective insecticides that will not disrupt biological control agents of BFSB. These experiments are currently underway.
These results from Bangladesh are similar to those from studies conducted in the Philippines in which event EE-1, the same event used to create the four Bt brinjal varieties used in these studies, was incorporated into open pollinated lines and provided almost complete control of BFSB in different locations over three cropping periods [ 10 ]. Furthermore, additional ecological studies in the Philippines [ 11 ] documented that many arthropod taxa are associated with Bt eggplants and their non-Bt comparators, but found few significant differences in seasonal mean densities of arthropod taxa between Bt and non-Bt eggplants when no insecticides were used. Principal Response Curve analyses showed no statistically significant impact of Bt eggplants on overall arthropod communities through time in any season. Furthermore, the Philippine studies found no significant adverse impacts of Bt eggplants on species abundance, diversity and community dynamics, particularly for beneficial NTOs. Similarly, in the present study we did not find any differences in the arthropod communities in any Bt brinjal variety compared to its non-Bt isoline. This is not surprising because the ecological effect of Cry1Ac has been extensively studied and shown to have little to no effect on non-target organisms outside of the Lepidoptera [ 12 – 24 ].
In most cases, statistically higher numbers of non-target pest arthropods were observed in no-spray plots irrespective of varieties, except for whitefly and mites. Furthermore, populations were similar in Bt and non-Bt isolines, irrespective of spray regime in most cases. Similar patterns have been observed before. In India, it has been reported that populations of major non-target insect pests (leafhoppers, whiteflies, ash weevils, aphids, dusky and red cotton bug, and green bug) and generalist predators (ladybirds, chrysopids, and spiders) did not differ significantly between Bt and non-Bt cotton lines, while their numbers were lower in insecticide protected than under unprotected conditions, except for aphids and whiteflies [ 25 ]. Ladybird beetles were more abundant in no-spray plots but similar abundances of non-target beneficial and other arthropods were observed in Bt and non-Bt isolines irrespective of spray regime in most cases. Other larger scale studies and meta-analyses have documented that Bt crops were much safer to non-target organisms than the alternative use of traditional insecticides for control of the pests targeted by the Bt proteins [ 17 , 19 , 21 , 26 ].
Plot sizes in this study were relatively small and the effect of this on study outcomes would depend somewhat on the general mobility of the species examined. While specific guidance on plot size for any given study is not available, there is general agreement that plots should be as large as possible to avoid inter-plot exchanges of arthropods [ 27 , 28 ]. In reality, plot size is typically dictated by experimental design issues, and space resources as they were here. Nonetheless, despite small plots sizes, this study clearly delineated the effects of Bt eggplant on target pest abundance and its associated impact on yield. Furthermore, other experiments and research syntheses that have examined a wide range of plots sizes have shown that plot size has a relatively small impact on the assessment of non-target effects [ 17 , 29 ]. Finally, the very small scale of individual farms growing eggplant in Bangladesh suggests that our studies are reflective of commercial practices, and thus of potential non-target effects.
Since their first introduction in 1996, biotech crops have been large scale field crops except for the relatively small-scale production of virus resistant papaya and squash and insect-resistant sweet corn, and the brief commercialization of Bt potatoes [ 3 ]. Bt brinjal is the first Bt vegetable crop introduced into a developing country and the results reported here indicate that it can be highly successful. An ex-ante study in 2005 estimated that the introduction of Bt eggplant into Bangladesh would result in a decrease of insecticides by 70–90%, increase yield by 15–30% and increase the gross return by 37–64% [ 30 ]. The data from this study supports these general predictions.
To realize these benefits of Bt brinjal for the long term, it is critical that EFSB does not rapidly evolve resistance to the Cry1Ac protein it produces. A government condition for the release of Bt brinjal in Bangladesh requires planting a refuge of non-Bt brinjal and training materials provided to farmers emphasizing that non-Bt brinjal should be planted around Bt fields as a structured refuge. In addition, to this refuge, there are many non-Bt varieties commonly grown in Bangladesh that also may serve as a natural refuge for resistance management. Nonetheless, it will be important to develop lines that express multiple Bt proteins because this is another key factor in delaying the evolution of resistance [ 31 ].
In conclusion, the four varieties of Bt eggplant examined here provide high levels of BFSB control, demonstrate higher gross returns than their non-Bt isolines and have the potential to greatly reduce insecticide inputs and their associated costs for management of this devastating pest. Additional controls of other pests in the system appear to be important and further development of management strategies for these will likely lead to further favorable economic returns in crop production for farmers growing brinjal in Bangladesh. In addition to providing excellent pest suppression, cultivation of Bt brinjal demonstrated no undesirable non-target effects on other arthropods in the system, especially those beneficial organisms that contribute important ecosystem services like biological control. Overall, careful stewardship will be critical to preserving this valuable pest control technology as adoption continues to increase from the more than 27,000 farmers who grew Bt brinjal in 2018 [ 32 ].
Acknowledgments
We gratefully acknowledge the support provided by the United States Agency for International Development (USAID) for their Feed the Future South Asia Eggplant Improvement Partnership (AID-OAA-A-15-00052). We are grateful for the help provided by the Bangladesh Agricultural Research Institute.
- 1. James C. Global status of commercialized biotech/GM crops: 2017. ISAAA Briefs, No 53, Ithaca, NY: ISAAA; 2013.
- 2. Brookes G, Barfoot P. GM crops: global socio-economic and environmental impacts 1996–2015. UK: PG economics LTD; 2017.
- View Article
- PubMed/NCBI
- Google Scholar
- 5. Karim ANMR. IPM for vegetable crops: issues and technology developments in Bangladesh. A keynote paper presented at the sixth biennial conference of Bangladesh Entomological Society held at BARI, Joydebpur, Gazipur, Bangladesh, October 7, 2004; 2004
- 6. Bangladesh Agricultural Research Institute (BARI). Integrated management for the brinjal shoot and fruit borer, Leucinodes orbonalis Guen. In: Annual Research Report,1993–94. Gazipur, Bangladesh: BARI;1994. pp. 94–96.
- 7. Shelton AM, Hokanson KE, Hautea DM, Hossain MJ, Hossain MA, Paranjape V, Hautea RA, McCandless L, Sarwer SH. Bt eggplant: A genetically engineered ‘minor’ crop comes of age in Bangladesh and the Philippines. ISB News Report. 2017; August 2017. https://vtechworks.lib.vt.edu/bitstream/handle/10919/78874/SHHHHPHMCS.pdf?sequence=1&isAllowed=y
- 8. Mondal MRI, Islam MS, Bhuiyan MAJ, Rahman MM, Alam MS, Rahman MHH. Vegetable crops. In: Hatboi KP, editor. Handbook on agro-technology, 5 th ed. Gazipur, Bangladesh: Bangladesh Agricultural Research Institute; 2011. pp 397–409.
- 9. Kovach J, Petzoldt C, Degni J, Tette J. A method to measure the environmental impact of pesticides. Geneva, NY: New York’s Food and Life Sciences Bulletin, NYSAES Cornell University; 1992. https://nysipm.cornell.edu/eiq
- 26. Naranjo SE. Effects of GM crops on non-target organisms. In: Ricroch A, Chopra S, Fleischer SJ, editors. Plant biotechnology: Experience and future prospects. New York: Springer; 2014. pp.129–142.
- 30. Islam SMF, Norton GW. 2007. Bt eggplant for fruit and shoot borer resistance in Bangladesh. In: Ramasamy C, Selvaraj KN, Norton GW, Vijayaraghavan K, editors. Economic and environmental benefits and costs of transgenic crops: Ex-ante assessment. Coimbatore, India, Tamil Nadu Agricultural University; 2007. pp. 91–106.
A Feasibility on the Establishment of Fusion of Eggplant Nuggets in Quirino Highway Novaliches Quezon City
- Michaella Dela Cruz
- Erdie Aguinaldo
- Glaiza Torres
- Eduardo Sereno
- Paul Vincent SD. Quinto, LPT
Many Filipinos love to eat vegetable, so the researchers created a simple eggplant nuggets for people especially for those people who don't eat vegetables. We created this dish to help those people who don't like to eat vegetables. The primary concept of the business is to produce quality eggplant nuggets that is healthy, tasty, affordable and within the budget. The product we produce has a unique quality, these product are for any people. This type of nuggets is most likely for kids, teens and also for adults. This is not just an ordinary tasting nuggets that is healthy but one that is also available at a very affordable price. The researchers chose survey because this method is easy to do. The researchers will just give free taste of their product to the respondents and allow them to answer the questions about the researchers' product or business. The survey result of our product (veggie nuggets) is 100% successful, because most of our respondents/customers like the taste of the product (Eggplant Nuggets), because of its new flavor, affordable price and nutritious benefits. Veggie Nuggets is much healthier compares to other foods because we add a very nutritious vegetable which is eggplant and carrots. The main focus of our business is to give the healthier version of veggie nuggets. Veggie Nuggets introduces new version of Eggplant nuggets made by the researchers which is to aim offer a new food not only delicious but also nutritious. According to Sally Kuzenchak, these veggie nuggets are healthy and simple to make and is perfect for lunch boxes and snacks. In the Philippines, we are able to have our own combination which fits our dishes. Researchers discover Veggie Nuggets and Eggplant Nuggets for those people who don't eat vegetables. Researchers created this dish to help those people who don't like to eat vegetables. The primary concept of the business is to produce quality Eggplant Nuggets that is healthy, tasty and affordable.

How to Cite
- Endnote/Zotero/Mendeley (RIS)
Most read articles by the same author(s)
- Pamela Besmonte, John Paul Candelario, Karl Paolo Colanggoy, Lourich Laroza, Cheremene Rose Villarmea, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Healthy Siomai (Vegetable Siomai) at Novaliches Bayan , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Joan L. Boñaga, John Reynald Arpon, Adriane Paul A. Datoon, James Ryan De Leon, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Product of “Puto Squash” in Glori Novaliches Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Ferlyn Misalang, Danica Jean Hibas, Ma. Katrin Balajadia, Danica Binban, Jessel Nuera, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Establishment of Ube Halaya Cheese Ballin Novaliches Bayan, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Mark Rafael Remontal, Kathereen Cortez, Jovie Lyca Damaso, Mariafe Samera, Jelly Ann Santos, Paul Vincent SD. Quinto, LPT, The Feasiblity Study on the Establishment of Veggie Siomai near Bestlink College of the Philippines (MV Campus) , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Ana Camila A. Pilar, Jomel M. Rivera, Kessiah P. Medrano, Ma. Chaelene Z. Lizada, John Owen I. Tolledo, Paul Vincent SD. Quinto, LPT, A Feasibility Study on Spicy Pastillas with Mango Jam Stuff in 1003 Qurino Highway, Sta.Monica Novaliches, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Honeyrose Martinez, Jeraldine Calumag, Sherlyn Carbonilla, Frances Joy Olival, Marian Fe Talinting, Jonaliza Perl Rey, Paul Vincent SD. Quinto, LPT, The Feasibility Study on the Establishment of "Veggie Empanada"near Glori Bayan, Novaliches, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Cheene Hanna L. Cepe, Cyrine S. Letada, April Joy Orante, Maribel Soriano, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Product of Cabbage Patty in Novaliches Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Rein Marc Cabaya, Jake Mateo, Ronalou Dangca, Mharco Armilla, Jojo Disono, Jojo Disono, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Establishment of Malunggay Patty Burgerin Barangay Gulod, Novaliches, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Michelle G. Morallos, Franz Kenneth Escribano, Christian Darryl Lloderiz, Joel Fontanilla, Paul Vincent SD. Quinto, LPT, A Feasibility Study on the Establishment of Fusion Carrots Puto in Novaliches, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
- Luzielle Ann Banda, Mary Ruth Dabal, Jenelyn Dalida, Keith Jasmine Garcia, Jeffrey Manzanilla, Paul Vincent SD. Quinto, LPT, A Feasibility Study on Bitter Melon Chocolate Chip Cookies in Glori Novaliches, Quezon City , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 1 No. 1 (2019): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.1, No.1, March 2019
Similar Articles
- Kenneth Bonita, Jaypee Cabig, Jehan Eslit, Rosita Marie Medina, Jaspher Orfelia, Alejandro Adovas, Enrollment System for Divine Trinity Learning Academy , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 2 No. 1 (2020): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.2, No1, March 2020
- Tyrone Catubao, Marc Ian Patrico, Austin Jones Exiomo, Florencio Mahidlawon, Crystal Jane Antonio, Alejandro Adovas, Scheduling System for Teacher of Lagro High School , Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research: Vol. 2 No. 1 (2020): Ascendens Asia Singapore – Bestlink College of the Philippines Journal of Multidisciplinary Research Abstracts, Vol.2, No1, March 2020
You may also start an advanced similarity search for this article.

Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Published: 14 May 2024
2023 summer warmth unparalleled over the past 2,000 years
- Jan Esper ORCID: orcid.org/0000-0003-3919-014X 1 , 2 ,
- Max Torbenson ORCID: orcid.org/0000-0003-2720-2238 1 &
- Ulf Büntgen 2 , 3 , 4
Nature ( 2024 ) Cite this article
912 Accesses
3320 Altmetric
Metrics details
We are providing an unedited version of this manuscript to give early access to its findings. Before final publication, the manuscript will undergo further editing. Please note there may be errors present which affect the content, and all legal disclaimers apply.
- Climate change
- Palaeoclimate
Including an exceptionally warm Northern Hemisphere (NH) summer 1 ,2 , 2023 has been reported as the hottest year on record 3-5 . Contextualizing recent anthropogenic warming against past natural variability is nontrivial, however, because the sparse 19 th century meteorological records tend to be too warm 6 . Here, we combine observed and reconstructed June-August (JJA) surface air temperatures to show that 2023 was the warmest NH extra-tropical summer over the past 2000 years exceeding the 95% confidence range of natural climate variability by more than half a degree Celsius. Comparison of the 2023 JJA warming against the coldest reconstructed summer in 536 CE reveals a maximum range of pre-Anthropocene-to-2023 temperatures of 3.93°C. Although 2023 is consistent with a greenhouse gases-induced warming trend 7 that is amplified by an unfolding El Niño event 8 , this extreme emphasizes the urgency to implement international agreements for carbon emission reduction.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
185,98 € per year
only 3,65 € per issue
Rent or buy this article
Prices vary by article type
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

The economic commitment of climate change

Climate extremes likely to drive land mammal extinction during next supercontinent assembly

Climate damage projections beyond annual temperature
Author information, authors and affiliations.
Department of Geography, Johannes Gutenberg University, Mainz, Germany
Jan Esper & Max Torbenson
Global Change Research Institute of the Czech Academy of Sciences, Brno, Czech Republic
Jan Esper & Ulf Büntgen
Department of Geography, University of Cambridge, Cambridge, United Kingdom
Ulf Büntgen
Department of Geography, Masaryk University, Brno, Czech Republic
You can also search for this author in PubMed Google Scholar
Corresponding author
Correspondence to Jan Esper .
Rights and permissions
Reprints and permissions
About this article
Cite this article.
Esper, J., Torbenson, M. & Büntgen, U. 2023 summer warmth unparalleled over the past 2,000 years. Nature (2024). https://doi.org/10.1038/s41586-024-07512-y
Download citation
Received : 16 January 2024
Accepted : 02 May 2024
Published : 14 May 2024
DOI : https://doi.org/10.1038/s41586-024-07512-y
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
By submitting a comment you agree to abide by our Terms and Community Guidelines . If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

Cities as Engines of Opportunities: Evidence from Brazil
Are developing-world cities engines of opportunities for low-wage earners? In this study, we track a cohort of young low-income workers in Brazil for thirteen years to explore the contribution of factors such as industrial structure and skill segregation on upward income mobility. We find that cities in the south of Brazil are more effective engines of upward mobility than cities in the north and that these differences appear to be primarily related to the exposure of unskilled workers to skilled co-workers, which in turn reflects industry composition and complexity. Our results suggest that the positive effects of urbanization depend on the skilled and unskilled working together, a form of integration that is more prevalent in the cities of southern Brazil than in northern cities. This segregation, which can decline with specialization and the division of labor, may hinder the ability of Brazil's northern cities to offer more opportunities for escaping poverty.
We acknowledge the support of Cristian Jara-Figueroa in the initial conceptualization of the empirical strategy. Barza and Viarengo gratefully acknowledges the financial support received from the Swiss National Science Foundation (Principal Investigator: Prof. Dr. Martina Viarengo; Research Grant n. 100018-176454). Hidalgo acknowledges the support of the Agence Nationale de la Recherche grant number ANR-19-P3IA-0004, the 101086712-LearnData-HORIZON-WIDERA-2022-TALENTS-01 financed by European Research Executive Agency (REA) (https://cordis.europa.eu/project/id/101086712), IAST funding from the French National Research Agency (ANR) under grant ANR-17-EURE-0010 (Investissements d'Avenir program), and the European Lighthouse of AI for Sustainability [grant number 101120237-HOR-IZON-CL4-2022-HUMAN-02]. The usual caveats apply. The views expressed herein are those of the authors and do not necessarily reflect the views of the National Bureau of Economic Research.
I have received speaking fees from organizations that organize members that invest in real estate markets, including the National Association of Real Estate Investment Managers, the Pension Real Estate Association and the Association for International Real Estate Investors.
MARC RIS BibTeΧ
Download Citation Data
Working Groups
Mentioned in the news, more from nber.
In addition to working papers , the NBER disseminates affiliates’ latest findings through a range of free periodicals — the NBER Reporter , the NBER Digest , the Bulletin on Retirement and Disability , the Bulletin on Health , and the Bulletin on Entrepreneurship — as well as online conference reports , video lectures , and interviews .

Are Markups Driving the Ups and Downs of Inflation?

Download PDF (158 KB)
FRBSF Economic Letter 2024-12 | May 13, 2024
How much impact have price markups for goods and services had on the recent surge and the subsequent decline of inflation? Since 2021, markups have risen substantially in a few industries such as motor vehicles and petroleum. However, aggregate markups—which are more relevant for overall inflation—have generally remained flat, in line with previous economic recoveries over the past three decades. These patterns suggest that markup fluctuations have not been a main driver of the ups and downs of inflation during the post-pandemic recovery.
In the recovery from the pandemic, U.S. inflation surged to a peak of over 7% in June 2022 and has since declined to 2.7% in March 2024, as measured by the 12-month change in the personal consumption expenditures (PCE) price index. What factors have been driving the ups and downs of inflation? Production costs are traditionally considered a main contributor, particularly costs stemming from fluctuations in demand for and supply of goods and services. As demand for their products rises, companies need to hire more workers and buy more intermediate goods, pushing up production costs. Supply chain disruptions can also push up the cost of production. Firms may pass on all or part of the cost increases to consumers by raising prices. Thus, an important theoretical linkage runs from cost increases to inflation. Likewise, decreases in costs should lead to disinflation.
Labor costs are an important factor of production costs and are often useful for gauging inflationary pressures. However, during the post-pandemic surge in inflation, nominal wages rose more slowly than prices, such that real labor costs were falling until early 2023. By contrast, disruptions to global supply chains pushed up intermediate goods costs, contributing to the surge in inflation (see, for example, Liu and Nguyen 2023). However, supply chains have more direct impacts on goods inflation than on services inflation, which also rose substantially.
In this Economic Letter , we consider another factor that might drive inflation fluctuations: changes in firms’ pricing power and markups. An increase in pricing power would be reflected in price-cost markups, leading to higher inflation; likewise, a decline in pricing power and markups could alleviate inflation pressures. We use industry-level measures of markups to trace their evolving impact on inflation during the current expansion. We find that markups rose substantially in some sectors, such as the motor vehicles industry. However, the aggregate markup across all sectors of the economy, which is more relevant for inflation, has stayed essentially flat during the post-pandemic recovery. This is broadly in line with patterns during previous business cycle recoveries. Overall, our analysis suggests that fluctuations in markups were not a main driver of the post-pandemic surge in inflation, nor of the recent disinflation that started in mid-2022.
Potential drivers of inflation: Production costs and markups
To support households and businesses during the pandemic, the Federal Reserve lowered the federal funds rate target to essentially zero, and the federal government provided large fiscal transfers and increased unemployment benefits. These policies boosted demand for goods and services, especially as the economy recovered from the depth of the pandemic.
The increase in overall demand, combined with supply shortages, boosted the costs of production, contributing to the surge in inflation during the post-pandemic recovery. Although labor costs account for a large part of firms’ total production costs, real labor costs were falling between early 2021 and mid-2022 such that the increases in prices outpaced those in nominal wages. This makes it unlikely that labor costs were driving the surge in inflation.
Instead, we focus on another potential alternative driver of inflation that resulted from firms’ ability to adjust prices, known as pricing power. As demand for goods surged early in the post-pandemic recovery, companies may have had a greater ability to raise their prices above their production costs, a gap known as markups. Following a sharp drop in spending at the height of the pandemic, people may have become eager to resume normal spending patterns and hence more tolerant to price increases than in the past. In fact, growth of nonfinancial corporate profits accelerated in the early part of the recovery (see Figure 1), suggesting that companies had increased pricing power. Some studies have pointed to the strong growth in nonfinancial corporate profits in 2021 as evidence that increased markups have contributed to inflation (see, for example, Weber and Wasmer 2023). However, the figure also shows that growth in corporate profits is typically volatile. Corporate profits tend to rise in the early stages of economic recoveries. Data for the current recovery show that the increase in corporate profits is not particularly pronounced compared with previous recoveries.
Figure 1 Profit growth for nonfinancial businesses
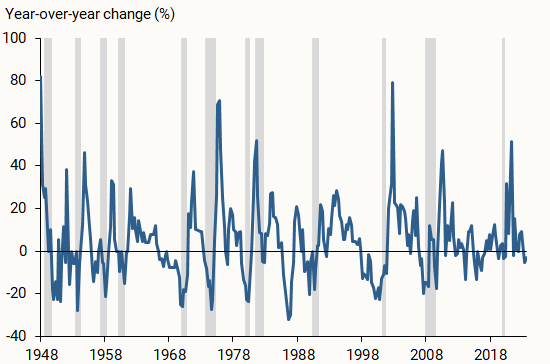
More importantly, corporate profits are an imperfect measure of a firm’s pricing power because several other factors can drive changes in profitability. For instance, much of the recent rise in corporate profits can be attributed to lower business taxes and higher subsidies from pandemic-related government support, as well as lower net interest payments due to monetary policy accommodation (Pallazzo 2023).
Instead of relying on profits as a measure of pricing power, we construct direct measures of markups based on standard economic models. Theory suggests that companies set prices as a markup over variable production costs, and that markup can be inferred from the share of a firm’s revenue spent on a given variable production factor, such as labor or intermediate goods. Over the period of data we use, we assume that the specific proportion of a company’s production costs going toward inputs does not change. If the share of a firm’s revenue used for inputs falls, it would imply a rise in the firm’s price-cost margin or markup. In our main analysis, we use industry-level data from the Bureau of Economic Analysis (BEA) to compute markups based on the share of revenue spent on intermediate inputs. Our results are similar if we instead use the share of revenue going toward labor costs.
We compare the evolution of markups to that of prices, as measured by the PCE price index, since the recovery from the pandemic. In constructing this price index, the BEA takes into account changes in product characteristics (for instance, size) that could otherwise bias the inflation measure by comparing the prices of inherently different products over time. Similarly, based upon standard economic theory, our markup measure implicitly captures changes in those characteristics (see, for example, Aghion et al. 2023).
The post-pandemic evolution of markups
We examine the evolution of markups in each industry since the third quarter of 2020, the start of the post-pandemic recovery. Figure 2 shows that some sectors, such as the motor vehicles and petroleum industries, experienced large cumulative increases in markups during the recovery. Markups also rose substantially in general merchandise, such as department stores, and for other services, such as repair and maintenance, personal care, and laundry services. Since the start of the expansion, markups in those industries rose by over 10%—comparable in size to the cumulative increases over the same period in the core PCE price index, which excludes volatile food and energy components. However, the surge in inflation through June 2022 was broad based, with prices also rising substantially outside of these sectors. Thus, understanding the importance of markups for driving inflation requires a macroeconomic perspective that examines the evolution of aggregate markups across all sectors of the economy.
Figure 2 Cumulative changes in markups for salient industries
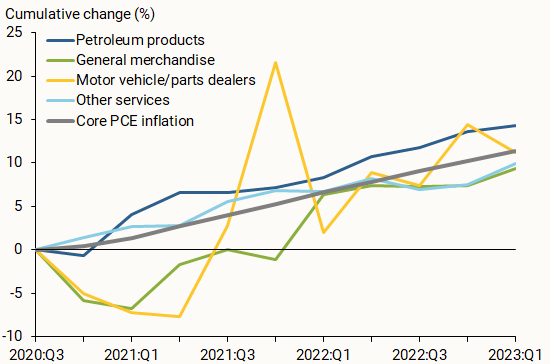
The role of aggregate markups in the economy
To assess how much markup changes contribute to movements in inflation more broadly, we use our industry-level measurements to calculate an aggregate markup at the macroeconomic level. We aggregate the cumulative changes in industry markups, applying two different weighting methods, as displayed in Figure 3. In the first method (green line), we match our industry categories to the spending categories in the core PCE price index for ease of comparison; we then use the PCE weights for each category to compute the aggregate markup. Alternatively, we use each industry’s cost weights to compute the aggregate markup (blue line). Regardless of the weighting method, Figure 3 shows that aggregate markups have stayed essentially flat since the start of the recovery, while the core PCE price index (gray line) rose by more than 10%. Thus, changes in markups are not likely to be the main driver of inflation during the recovery, which aligns with results from Glover, Mustre-del-Río, and von Ende-Becker (2023) and Hornstein (2023) using different methodologies or data. Markups also have not played much of a role in the slowing of inflation since the summer of 2022.
Figure 3 Cumulative changes in aggregate markups and prices
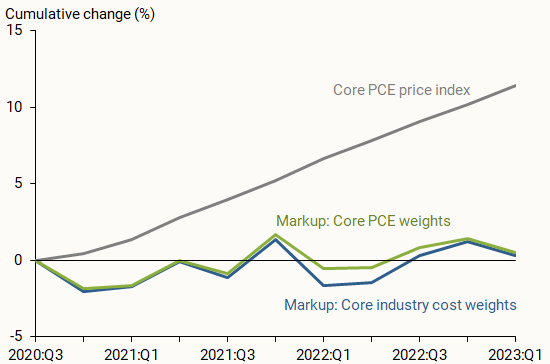
Moreover, the path of aggregate markups over the past three years is not unusual compared with previous recoveries. Figure 4 shows the cumulative changes in aggregate markups since the start of the current recovery (dark blue line), alongside aggregate markups following the 1991 (green line), 2001 (yellow line), and 2008 (light blue line) recessions. Aggregate markups have stayed roughly constant throughout all four recoveries.
Figure 4 Cumulative changes of aggregate markups in recoveries
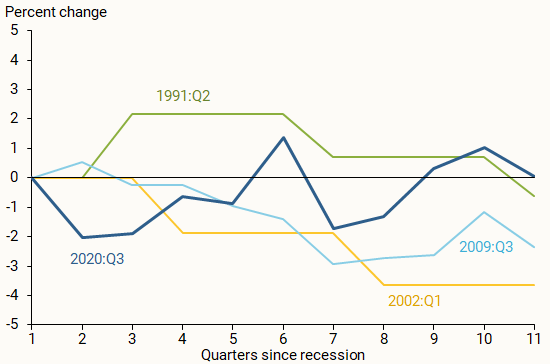
Firms’ pricing power may change over time, resulting in markup fluctuations. In this Letter , we examine whether increases in markups played an important role during the inflation surge between early 2021 and mid-2022 and if declines in markups have contributed to disinflation since then. Using industry-level data, we show that markups did rise substantially in a few important sectors, such as motor vehicles and petroleum products. However, aggregate markups—the more relevant measure for overall inflation—have stayed essentially flat since the start of the recovery. As such, rising markups have not been a main driver of the recent surge and subsequent decline in inflation during the current recovery.
Aghion, Philippe, Antonin Bergeaud, Timo Boppart, Peter J. Klenow, and Huiyu Li. 2023. “A Theory of Falling Growth and Rising Rents.” Review of Economic Studies 90(6), pp.2,675-2,702.
Glover, Andrew, José Mustre-del-Río, and Alice von Ende-Becker. 2023. “ How Much Have Record Corporate Profits Contributed to Recent Inflation? ” FRB Kansas City Economic Review 108(1).
Hornstein, Andreas. 2023. “ Profits and Inflation in the Time of Covid .” FRB Richmond Economic Brief 23-38 (November).
Liu, Zheng, and Thuy Lan Nguyen. 2023. “ Global Supply Chain Pressures and U.S. Inflation .” FRBSF Economic Letter 2023-14 (June 20).
Palazzo, Berardino. 2023. “ Corporate Profits in the Aftermath of COVID-19 .” FEDS Notes , Federal Reserve Board of Governors, September 8.
Weber, Isabella M. and Evan Wasner. 2023. “Sellers’ Inflation, Profits and Conflict: Why Can Large Firms Hike Prices in an Emergency?” Review of Keynesian Economics 11(2), pp. 183-213.
Opinions expressed in FRBSF Economic Letter do not necessarily reflect the views of the management of the Federal Reserve Bank of San Francisco or of the Board of Governors of the Federal Reserve System. This publication is edited by Anita Todd and Karen Barnes. Permission to reprint portions of articles or whole articles must be obtained in writing. Please send editorial comments and requests for reprint permission to [email protected]
Haifeng Xu Wins Best Paper Award at Leading AI Conference for Pioneering Research on Mechanism Design for LLMs
As this year’s Web Conference is under way, pioneering research work by Assistant Professor of Computer Science and Data Science Haifeng Xu and his collaborators has been announced as the winner for their prestigious Best Paper Award.

Xu’s paper, entitled “ Mechanism Design for Large Language Models ,” was selected from amongst 2008 submissions.
This paper lays out a newly developed method to aggregate language generations from multiple self-interested LLM agents into a single text generation. It does so by accounting for these LLM agents’ self-interests in an incentive-compatible way. As summarized in the meta review, “the review team unanimously finds the paper novel, well-executed, and … has potential to be a landmark paper sparking a new line of research linking LLMs and mechanism design.”
This paper is a joint work with Google Researchers. The technology Xu and his team developed has been tested on Google’s LLM model Bard and Xu reports that it performs very well. According to Xu, the nice (and often very rare) combination of both strong theoretical development and real-world implementation on Bard is probably a key reason for the paper to be named the Best Paper.
Congratulations, Haifeng!
This article was originally published by the Data Science Institute.
Related News

Fred Chong Receives Quantrell Award for Excellence in Teaching

Unveiling Attention Receipts: Tangible Reflections on Digital Consumption

NASA to Launch UChicago Undergraduates’ Satellite

University of Chicago Computer Science Researchers To Present Ten Papers at CHI 2024

Two UChicago MPCS Students Win the Apple Swift Student Challenge

How Artificial Intelligence Can Transform U.S. Energy Infrastructure

Community Data Fellow Stephania Tello Zamudio helps broaden internet access for Illinois residents

Two UChicago CS Students Awarded NSF Graduate Research Fellowship

Non-Unital Noise Adds a New Wrinkle to the Quantum Supremacy Debate

The Science of Computer Security: An Interview with Grant Ho, Assistant Professor in Computer Science

Four Students Receive Honorable Mention in CRA Undergraduate Research Awards

Navigating the Intersection of Technology and Public Policy: The Journey of Ranya Sharma at UChicago
Suggestions or feedback?
MIT News | Massachusetts Institute of Technology
- Machine learning
- Social justice
- Black holes
- Classes and programs
Departments
- Aeronautics and Astronautics
- Brain and Cognitive Sciences
- Architecture
- Political Science
- Mechanical Engineering
Centers, Labs, & Programs
- Abdul Latif Jameel Poverty Action Lab (J-PAL)
- Picower Institute for Learning and Memory
- Lincoln Laboratory
- School of Architecture + Planning
- School of Engineering
- School of Humanities, Arts, and Social Sciences
- Sloan School of Management
- School of Science
- MIT Schwarzman College of Computing
Using ideas from game theory to improve the reliability of language models
Press contact :.

Previous image Next image
Imagine you and a friend are playing a game where your goal is to communicate secret messages to each other using only cryptic sentences. Your friend's job is to guess the secret message behind your sentences. Sometimes, you give clues directly, and other times, your friend has to guess the message by asking yes-or-no questions about the clues you've given. The challenge is that both of you want to make sure you're understanding each other correctly and agreeing on the secret message.
MIT Computer Science and Artificial Intelligence Laboratory (CSAIL) researchers have created a similar "game" to help improve how AI understands and generates text. It is known as a “consensus game” and it involves two parts of an AI system — one part tries to generate sentences (like giving clues), and the other part tries to understand and evaluate those sentences (like guessing the secret message).
The researchers discovered that by treating this interaction as a game, where both parts of the AI work together under specific rules to agree on the right message, they could significantly improve the AI's ability to give correct and coherent answers to questions. They tested this new game-like approach on a variety of tasks, such as reading comprehension, solving math problems, and carrying on conversations, and found that it helped the AI perform better across the board.
Traditionally, large language models answer one of two ways: generating answers directly from the model (generative querying) or using the model to score a set of predefined answers (discriminative querying), which can lead to differing and sometimes incompatible results. With the generative approach, "Who is the president of the United States?" might yield a straightforward answer like "Joe Biden." However, a discriminative query could incorrectly dispute this fact when evaluating the same answer, such as "Barack Obama."
So, how do we reconcile mutually incompatible scoring procedures to achieve coherent, efficient predictions?
"Imagine a new way to help language models understand and generate text, like a game. We've developed a training-free, game-theoretic method that treats the whole process as a complex game of clues and signals, where a generator tries to send the right message to a discriminator using natural language. Instead of chess pieces, they're using words and sentences," says Athul Jacob, an MIT PhD student in electrical engineering and computer science and CSAIL affiliate. "Our way to navigate this game is finding the 'approximate equilibria,' leading to a new decoding algorithm called 'equilibrium ranking.' It's a pretty exciting demonstration of how bringing game-theoretic strategies into the mix can tackle some big challenges in making language models more reliable and consistent."
When tested across many tasks, like reading comprehension, commonsense reasoning, math problem-solving, and dialogue, the team's algorithm consistently improved how well these models performed. Using the ER algorithm with the LLaMA-7B model even outshone the results from much larger models. "Given that they are already competitive, that people have been working on it for a while, but the level of improvements we saw being able to outperform a model that's 10 times the size was a pleasant surprise," says Jacob.
"Diplomacy," a strategic board game set in pre-World War I Europe, where players negotiate alliances, betray friends, and conquer territories without the use of dice — relying purely on skill, strategy, and interpersonal manipulation — recently had a second coming. In November 2022, computer scientists, including Jacob, developed “Cicero,” an AI agent that achieves human-level capabilities in the mixed-motive seven-player game, which requires the same aforementioned skills, but with natural language. The math behind this partially inspired the Consensus Game.
While the history of AI agents long predates when OpenAI's software entered the chat in November 2022, it's well documented that they can still cosplay as your well-meaning, yet pathological friend.
The consensus game system reaches equilibrium as an agreement, ensuring accuracy and fidelity to the model's original insights. To achieve this, the method iteratively adjusts the interactions between the generative and discriminative components until they reach a consensus on an answer that accurately reflects reality and aligns with their initial beliefs. This approach effectively bridges the gap between the two querying methods.
In practice, implementing the consensus game approach to language model querying, especially for question-answering tasks, does involve significant computational challenges. For example, when using datasets like MMLU, which have thousands of questions and multiple-choice answers, the model must apply the mechanism to each query. Then, it must reach a consensus between the generative and discriminative components for every question and its possible answers.
The system did struggle with a grade school right of passage: math word problems. It couldn't generate wrong answers, which is a critical component of understanding the process of coming up with the right one.
“The last few years have seen really impressive progress in both strategic decision-making and language generation from AI systems, but we’re just starting to figure out how to put the two together. Equilibrium ranking is a first step in this direction, but I think there’s a lot we’ll be able to do to scale this up to more complex problems,” says Jacob.
An avenue of future work involves enhancing the base model by integrating the outputs of the current method. This is particularly promising since it can yield more factual and consistent answers across various tasks, including factuality and open-ended generation. The potential for such a method to significantly improve the base model's performance is high, which could result in more reliable and factual outputs from ChatGPT and similar language models that people use daily.
"Even though modern language models, such as ChatGPT and Gemini, have led to solving various tasks through chat interfaces, the statistical decoding process that generates a response from such models has remained unchanged for decades," says Google Research Scientist Ahmad Beirami, who was not involved in the work. "The proposal by the MIT researchers is an innovative game-theoretic framework for decoding from language models through solving the equilibrium of a consensus game. The significant performance gains reported in the research paper are promising, opening the door to a potential paradigm shift in language model decoding that may fuel a flurry of new applications."
Jacob wrote the paper with MIT-IBM Watson Lab researcher Yikang Shen and MIT Department of Electrical Engineering and Computer Science assistant professors Gabriele Farina and Jacob Andreas, who is also a CSAIL member. They presented their work at the International Conference on Learning Representations (ICLR) earlier this month, where it was highlighted as a "spotlight paper." The research also received a “best paper award” at the NeurIPS R0-FoMo Workshop in December 2023.
Share this news article on:
Press mentions, quanta magazine.
MIT researchers have developed a new procedure that uses game theory to improve the accuracy and consistency of large language models (LLMs), reports Steve Nadis for Quanta Magazine . “The new work, which uses games to improve AI, stands in contrast to past approaches, which measured an AI program’s success via its mastery of games,” explains Nadis.
Previous item Next item
Related Links
- Article: "Game Theory Can Make AI More Correct and Efficient"
- Jacob Andreas
- Athul Paul Jacob
- Language & Intelligence @ MIT
- Computer Science and Artificial Intelligence Laboratory (CSAIL)
- Department of Electrical Engineering and Computer Science
- MIT-IBM Watson AI Lab
Related Topics
- Computer science and technology
- Artificial intelligence
- Human-computer interaction
- Natural language processing
- Game theory
- Electrical Engineering & Computer Science (eecs)
Related Articles

Reasoning and reliability in AI

Explained: Generative AI

Synthetic imagery sets new bar in AI training efficiency

Simulating discrimination in virtual reality
More mit news.

Janabel Xia: Algorithms, dance rhythms, and the drive to succeed
Read full story →

Jonathan Byrnes, MIT Center for Transportation and Logistics senior lecturer and visionary in supply chain management, dies at 75

Researchers develop a detector for continuously monitoring toxic gases

The beauty of biology

Navigating longevity with industry leaders at MIT AgeLab PLAN Forum

Jeong Min Park earns 2024 Schmidt Science Fellowship
- More news on MIT News homepage →
Massachusetts Institute of Technology 77 Massachusetts Avenue, Cambridge, MA, USA
- Map (opens in new window)
- Events (opens in new window)
- People (opens in new window)
- Careers (opens in new window)
- Accessibility
- Social Media Hub
- MIT on Facebook
- MIT on YouTube
- MIT on Instagram
IMAGES
VIDEO
COMMENTS
Eggplant (Solanum melongena L.) is an increasingly popular crop in the United States. In the southeastern United States, eggplant is often produced with high levels of irrigation water [above the rate of crop evapotranspiration (ETc)], resulting in water waste and nitrogen (N) leaching. The objective of this research was to assess the effects of irrigation rate on plant growth and fruit yield ...
This paper targeted to investigate rehydration characteristics of dehydrated black beauty eggplant (which was dried under sun, by hot air, microwave, and infrared) at 25, 50, and 75°C, and to ...
Introduction. Brinjal eggplant (Solanum melongena L.) is a warm-weather crop mostly cultivated in tropical and subtropical regions of the world.Two other cultivated eggplant species, the scarlet eggplant (S. aethiopicum L.) and the gboma eggplants (S. macrocarpon L.), are less known but have local importance in sub-Saharan Africa (Schippers, 2000; Daunay and Hazra, 2012).
Well-drained sandy loam, loa m, or clay loam soils having a good supply of or ganic matter a nd pH of. 6.0- 6.5 are best for growing eggplants. Plant spacing var ies from 45 to 60 cm between ...
Eggplant is one of the most important vegetable crops known for its nutritive benefits due to the abundance of various bioactive compounds, which include proteins, vitamins, minerals, carbohydrates, phenolics, and dry matter content. ... Feature papers represent the most advanced research with significant potential for high impact in the field ...
Antioxidants. Phenols. Eggplant is a vegetable crop that is grown around the world and can provide significant nutritive benefits thanks to its abundance of vitamins, phenolics and antioxidants. In addition, eggplant has potential pharmaceutical uses that are just now becoming recognized. As compared to other crops in the ….
Eggplant, Solanum melongena L. (2n = 24)—also known as aubergine or brinjal—is an important solanaceous vegetable crop in many countries. It is a native of India. Eggplant is a good source of minerals and vitamins, and in total nutritional value, it can be compared with tomato. The important eggplant-growing countries are India, Japan ...
Eggplant is rich in saccharides, such as fructose (13750 mg/kg), glucose (13270 mg/kg) and, to a lesser extent sucrose (5030 mg/kg) ( Ayaz et al., 2015 ). S. melongena also contains inositol and its derivatives which have positive effects on human health, including the treatment of insulin resistance ( Michell, 2007 ).
Eggplant from young plants grew faster and ... competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. ... y Tecnológica (PICT 2015-3690) for financial support. LV and MD are fellows of CONICET. MJZ, ARV, MLL and AC are research members of CONICET (Consejo Nacional de ...
1.3.1 Eggplant as a Model for Parallel Evolution. As mentioned above, the domestication of multiple members of the Solanaceae has been used as a model to study convergent evolution. During domestication, human selection on fruit colour, taste, shape and size has been pervasive across many crops (Meyer et al. 2012a), and in grass crops, some traits are controlled by orthologous genes, for ...
The quality of the eggplant genome assembly was further assessed (Supplementary Fig. S1).The alignment rate of all short reads to the genome was ~99.48%, covering 91.24% of the genome.
Eggplants grow into 98 different species with different shapes and colours. They reduce the risk of cancer, cardio vascular diseases, pre-menstrual syndrome, amenorrhea, ante natal anaemia and ...
Effect of Relay Intercropping with Normal Garlic or Green Garlic on POD Activity in Eggplant Leaves. As shown in Figure 2B, the POD activity in 2011 initially increased and then decreased, whereas in 2012, it overall continued to increase.In 2011, the POD activity in the NG treatment was lower than that in the CK treatment for all sampling dates, and most of the differences were significant.
Introduction. Brinjal eggplant (Solanum melongena L.) is a warm-weather crop mostly cultivated in tropical and subtropical regions of the world.Two other cultivated eggplant species, the scarlet eggplant (S. aethiopicum L.) and the gboma eggplants (S. macrocarpon L.), are less known but have local importance in sub-Saharan Africa (Schippers, 2000; Daunay and Hazra, 2012).
Eggplant is the fifth economically most important vegetable in the Solanaceae family after tomato, potato, chili, and tobacco. ... provides an outlook for future research directions and describes possible research applications. Feature papers are submitted upon individual invitation or recommendation by the scientific editors and must receive ...
Results of the studies presented in this paper indicate that Bt eggplant OP lines expressed the Cry1Ac protein in relevant plant parts primarily attacked by EFSB at the appropriate growth stages throughout the productive life of the crop. ... This research was funded through the United States Agency for International Development (USAID ...
eggplant by keep preserving the environment. MATERIALS AND METHODS The research was conducted at the field of andosol in Poncokusumo - Malang, 600 m asl, pH 5.4, from August untuil December 2013. Materials of the research are tray as the seedbed and strong green variety of eggplant seed. The applicable fertilizers include goat manure,
Eggplant or brinjal (Solanum melongena) is a popular vegetable grown throughout Asia where it is attacked by brinjal fruit and shoot borer (BFSB) (Leucinodes orbonalis). Yield losses in Bangladesh have been reported up to 86% and farmers rely primarily on frequent insecticide applications to reduce injury. Bangladesh has developed and released four brinjal varieties producing Cry1Ac (Bt ...
conducted on eggplant at cultivating plot. A total of four- furrowed experimental plots were established manually. Each furrow was 5.48 m long and 0.61 m wide, with 0.91 m in between furrows. Each furrow contained six seedlings and the space between seedlings was 0.91 m. Manual elimination of weed control was done daily throughout research period.
Many Filipinos love to eat vegetable, so the researchers created a simple eggplant nuggets for people especially for those people who don't eat vegetables. We created this dish to help those people who don't like to eat vegetables. The primary concept of the business is to produce quality eggplant nuggets that is healthy, tasty, affordable and within the budget.
Here, we combine observed and reconstructed June-August (JJA) surface air temperatures to show that 2023 was the warmest NH extra-tropical summer over the past 2000 years exceeding the 95% ...
PDF | Eggplant is a vegetable crop where its growth rate and yield quality are largely influenced by fertilizer application. The application of organic... | Find, read and cite all the research ...
In our model of the streetlight effect, we predict that when data shines a light on attractive but not optimal projects, it can severely narrow the breadth of exploration and lower individual and group payoffs. We test our predictions in an online lab experiment and show that the availability of data on the true value of one project can lower ...
1. Introduction. Eggplant (Solanum melongena) is a vegetable belonging to the Nightshade family [1], and it is the third most relevant crop behind potatoes and tomatoes [2].The demand for eggplant has increased due to its high nutritional value, mainly due to its high fiber value [3, 4] and its important content of phenols with antioxidant activity [5].
Working Paper 32426. DOI 10.3386/w32426. Issue Date May 2024. Are developing-world cities engines of opportunities for low-wage earners? In this study, we track a cohort of young low-income workers in Brazil for thirteen years to explore the contribution of factors such as industrial structure and skill segregation on upward income mobility. We ...
These patterns suggest that markup fluctuations have not been a main driver of the ups and downs of inflation during the post-pandemic recovery. In the recovery from the pandemic, U.S. inflation surged to a peak of over 7% in June 2022 and has since declined to 2.7% in March 2024, as measured by the 12-month change in the personal consumption ...
As this year's Web Conference is under way, pioneering research work by Assistant Professor of Computer Science and Data Science Haifeng Xu and his collaborators has been announced as the winner for their prestigious Best Paper Award.. The Web Conference is a premier international conference on AI, Information Retrieval, and Web Technology. Since 1989, the Web Conference has focused on the ...
MIT researchers' "consensus game" is a game-theoretic approach for language model decoding. The equilibrium-ranking algorithm harmonizes generative and discriminative querying to enhance prediction accuracy across various tasks, outperforming larger models and demonstrating the potential of game theory in improving language model consistency and truthfulness.
May 14, 2024 8:00 am ET. Text. Fake studies have flooded the publishers of top scientific journals, leading to thousands of retractions and millions of dollars in lost revenue. The biggest hit has ...
The study was aimed to investigate the. eggplant production (y ield and quality) under urban conditions in. substrate culture (safety and economic). The number of total fruit for. each plant was ...